主题:【原创】死亡起源 The Origin of Death -- az09
家园 错字有点多了
家园 死亡起源(十三)—— 细胞的主动自杀(凋亡) 续上,死亡起源(十二)
这篇文章我一直希望用一种科普的手法来写,只是生命中的许多机制实在太复杂,很难用简单的文字来描述。我知道,如果各位看官有耐心看到这里,就会发现,这篇文章的某些部分已经开始变得有些象教科书一般的枯燥了,呵呵。不过,这篇文章中最枯燥的部分已经快过去了,下面稍微复杂一些的,是细胞的自杀(凋亡),不过,如果看过前面部分关于“受体”和“激素”的介绍以后,其实也不难理解。细胞作为我们生命中的基本单位,它们的主动自杀行为,也是本文不可绕过的部分。
3.2.6 细胞的主动自杀 ——凋亡(apoptosis)
细胞作为我们生命的一个基本单位,其实有点像一个计算机程序设计中被封装的“对象”(Object)。我们本就是从单细胞生物演化过来的,所以,这个基本单元在亿万年的演化过程中,早已演化出了它自己的一套非常独立而且被封装的机制。和计算机程序中被封装的各种“对象”或者"函数”一样,它也拥有“应用程序接口”,可以接受各种“输入参数”和“输入信号”,并对这些信号做出各种反应。它自身内部也有自己的一套独立的控制程序,有自己独立的生命控制循环。细胞各种复杂行为当中,和我们相关的话题是:它在 “永生”的同时,也是在不断的“自杀”(凋亡)的。而且,相比起细胞坏死,细胞的凋亡,是更常见的细胞死亡形式。
细胞也是会死亡的。细胞的死亡分为两种:1.细胞坏死(Necrosis);2.凋亡(apoptosis)。
细胞坏死(Necrosis)指的是细胞或活体组织因为非正常原因造成的永久性死亡。这通常是指因外在因素造成非正常、病理性的细胞死亡,它会造成疾病,甚至个体死亡。
相对于细胞坏死,我们体内更常见的细胞死亡方式是“细胞的凋亡”(apoptosis),这是一种细胞主动的程序化自杀行为(programmed cell death)。而且,和细胞坏死不同的是,细胞的凋亡,不会引发机体的“炎症反应”。(注:“炎症反应“(Inflammation)是指我们的免疫系统中的“先天免疫系统”为移除有害刺激或病源体及促进修复的保护措施,并非如“后天免疫系统”般,是针对特定病源体。关于“炎症反应”,我们在讨论癌症的时候,还会提到。)
细胞的凋亡分为“内源性凋亡”和“外源性凋亡”。简单的说起来,细胞就像是一个计算机程序中被封装的“对象”,当它接受到外界通知它死亡的信号的时候,它通过一系列复杂的级联反应,自己将自己破坏,导致死亡。同时,作为一个封装对象,它内部也有一套自杀程序。当它感应到自身内部出现了一些问题的时候(比如DNA遭到破坏),也可能触发它启动它的自杀程序,导致它的死亡。它最后会分裂成许多个被细胞膜包裹的“凋亡小体”(apoptotic bodies),然后被巨噬细胞吞噬。由于整个过程中,细胞器(organelle)等细胞内容物并没有泄漏溢出,所以细胞的凋亡不会引发炎症反应。(注:细胞遭到破坏后,内容物溢出会刺激免疫系统中的“先天免疫系统”,产生红肿等炎症反应)
关于细胞凋亡,稍微详细一些的过程是这样的:
1. 外源途径:我们前面已经讨论过了,细胞表面是有许多“受体”(Receptor) 的,这些受体的作用,就是用来和“配体”(ligand)结合,接受外部给它的信号的。这个配体可以是可以是任何的小分子或离子甚至是蛋白质。这和计算机程序中,各种“封装对象”的参数及其输入方式是一模一样的。细胞的众多受体中,有些受体是所谓的“死亡受体”(death receptors),它们用来接收的,便是“死亡信号”。如下图所示,细胞表面上的FAS,TNF-R1,TRAIL-R1等等,都是“死亡受体”或者 “肿瘤坏死因子受体”,它们与配体结合,可以对细胞产生死亡激励信号,激活细胞内部的“凋亡蛋白酶”(Caspase)等等,再通过一系列的级联反应,最后导致细胞的凋亡。
2. 内源途径(也叫线粒体途径):细胞作为一个独立的封装对象,除了接受外部信号外,它也可以感知自己本身出现的问题。比如当它的DNA受到损伤时,这套自杀机制也可能会被激发。它通过一系列程序,改变线粒体内外膜的通透性,使得线粒体内部的一些内容物(如细胞色素C等等)释放到细胞质中,然后这些物质也会激活“凋亡蛋白酶”,再通过一系列的级联反应,最后导致细胞的凋亡。大体就是这么个过程。
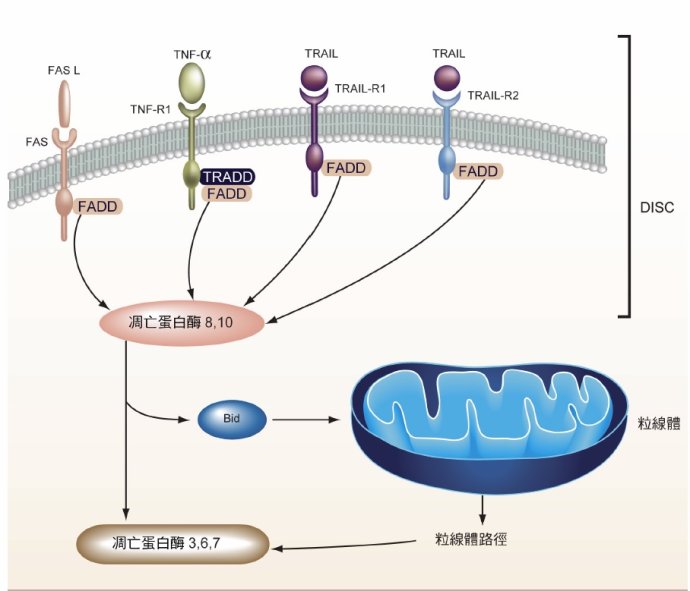
图55. 细胞主动自杀(凋亡)的两种信号通道。(1)外源途径,图中可以看见细胞膜上面的诸如FAS,TNF-R1,TRAIL-R1等等受体,这些受体是所谓的“死亡受体”或者“肿瘤坏死因子受体”,当它们接受到“死亡信号”,也就是那些小分子的“配体”(图中的FAS L, TNF, TRAIL等等)信号,会激活凋亡蛋白,导致细胞的凋亡。(2)内源途径,通常由线粒体触发,也会激活凋亡蛋白,导致细胞凋亡。
 图56. 组织中细胞凋亡过程示意。其实细胞的凋亡,未必是因为不健康,有些时候,仅仅是结构或者功能的需要,我们的机体也会诱导我们组织的某些细胞凋亡,特别在胚胎发育过程中,这种被诱导的细胞凋亡非常普遍。比如:胚胎发育中,眼睛的玻璃体和晶状体的细胞凋亡,是实现眼睛对光线的通透的重要的一步。 图片来自维基百科
图56. 组织中细胞凋亡过程示意。其实细胞的凋亡,未必是因为不健康,有些时候,仅仅是结构或者功能的需要,我们的机体也会诱导我们组织的某些细胞凋亡,特别在胚胎发育过程中,这种被诱导的细胞凋亡非常普遍。比如:胚胎发育中,眼睛的玻璃体和晶状体的细胞凋亡,是实现眼睛对光线的通透的重要的一步。 图片来自维基百科花上这么一小段篇幅介绍细胞的凋亡,主要是为了方便后面的讨论。总之,我们知道了,我们的细胞是浸泡在一个信号的海洋里面的,诸多信号中,有一类信号是“死亡信号”,我们的细胞可以接收“死亡信号”,并在“死亡信号”的通知下,主动自杀死亡。有这么个概念就足够了。
同时,相信大家都应该知道,我们体内的细胞是一直在死亡,一直在更新的,只是,我们体内的细胞的死亡,主要是因为细胞的主动自杀,而不仅仅是所谓的细胞的“老死”。
关于细胞的凋亡机制实际上非常复杂,同样也存在着许多未了机制,不过,我们暂时把细胞当作一个可以接受信号和参数的,和计算机程序设计中类似的,被封装的“对象”来看,也就够了………
待续..........请点击: 死亡起源(十四)
通宝推:铁手,唵啊吽,河区分,浣花岛主,明心灵竹,pattern,北纬42度,乔治·奥威尔,迷惑不解,石狼,喜你,李根,家园 死亡起源(十二)—— 寿命调节中的反馈机制 续上,死亡起源(十一)
比线虫更复杂的,是哺乳动物在饥饿压力下的反应。其实饥饿可以导致寿命延长本就是在作为哺乳动物的老鼠的身上首先被观察到的。
2012年10月得克萨斯大学西南医疗中心(UT Southwestern Medical Center)的研究者们《eLife》杂志上发表题为《饥饿激素,成纤维细胞生长因子21(FGF-21),延长小鼠寿命》[23]的研究论文。ScienceDaily 为此还专门配发了一篇评论:《饥饿激素显著延长小鼠寿命而无需限食》 [24] 。关于Fibroblast Growth Factor-21(FGF-21)的研究是最近比较热门的一项研究,因为它和许多有趣的生命控制机制都相关。FGF-21是一种所谓的“饥饿激素”,它主要由肝脏分泌,在空腹时,肝脏会分泌这种激素以此让身体做出适应饥饿的反应,它是一种行为很像激素的生长因子。人体的其他部位包括下丘脑也会分泌这种物质。得克萨斯大学的研究者们冒着失败的风险,做了一个很昂贵的实验。他们花了几年的时间,培养了一些可以过分表达和分泌FGF-21的转基因小鼠,并观察他们的寿命。他们并没有对这些小鼠限制食物,而是提供了足够的食物,不过观察的结果是,这些FGF-21长期过量分泌的小鼠的平均寿命要比普通小鼠增加30%,特别是对于雌鼠来说,其寿命要增加40%之多!研究者对此很兴奋,该文章的共同作者、他们的药理学主席David Mangelsdorf博士说:“长期过量生成FGF21激素使得小鼠无需减少食物摄取即可获得极长的寿命。它在没有节食的情况下模拟并获得了需要通过节食才能获得的健康与益处。”
不过这次我的视点还是没有集中到小鼠的寿命的延长上,因为这我看来,这本就天经地义。我视点在FGF-21的副作用上。因为包括该文的研究者也承认,FGF-21过量分泌的小鼠要比普通的野生小鼠体型更瘦小,并且伴有严重的骨质疏松。另外,最关键的一点是,FGF-21会导致雌鼠不育! 我们只要比较一下前面提到的线虫为什么延长了寿命,再和这个例子里面小鼠做一个对比,就可以观察到一个共同点——它们都抑制了生殖系统。
2013年8月,《Nature》的子刊《Nature Medicine》刊登了一篇文章“《FGF-21有助于神经内分泌控制雌性的生殖》(FGF21 contributes to neuroendocrine control of female reproduction)[25],描述了过量分泌的FGF-21是如何抑制雌性小鼠的生殖系统的。文章指出,FGF-21可以抑制雌性小鼠的生殖系统,导致性腺机能减退,以及雌激素等激素的降低。而且有趣的是,FGF-21在导致雌鼠不育的同时,却也使得雌鼠获得超过雄鼠的寿命的延长。雌鼠获得的寿命的延长要比平均寿命延长高10%,达到惊人的40%。
另外,除了导致雌鼠不育以外,“饥饿激素”FGF-21会导致食物和营养并不短缺的小鼠出现严重的骨质疏松也很有趣。其实生命本就有许多类似的抑制机制。比如,当我们遇到危险逃命的时候,我们的交感神经会变得十分兴奋,而副交感神经则受到压抑,表现出来的就是与逃命相关的项目获得增强,比如心跳加快,肾上腺素大量分泌,而与迫在眉睫的危险无关的,比如生殖、胃的消化、唾液的分泌等等,都会受到抑制,体内对这些部分提供的血液也会随之减少。这本就是生物的一个非常正常的应急反应——把能量用在最需要的地方。关于骨质疏松症,我解释一下:我们体内的钙循环是和骨骼相关的,我们的骨骼即便是在成年以后也是非常活跃的,一直在经历着生长和破坏的过程,也在不停的在与身体进行钙离子的循环和交换。钙离子对于身体来说极为重要,它是机体各项生理活动不可缺少的离子。 它对于维持细胞膜两侧的生物电位,维持正常的神经传导功能,维持正常的肌肉伸缩与舒张功能以及神经——肌肉传导功能都至关重要。而我们的骨骼,正是我们身体钙离子的储藏库。正常情况下,骨骼和其他器官之间的钙离子的循环会处于一个平衡的状态。不过随着年龄的增大,在某种机制的作用下(表现之一是老年人对钙的吸收率快速下降),钙离子的循环会变得不平衡,所以常常说老年人要补钙。另外,怀孕期的妇女因为母体需要将大量的钙提供给胎儿建造身体和骨骼,也会导致母体临时性的缺钙和骨质疏松——也就是说,母体把自己库存的钙大量提供给了胎儿,导致了自己体内的钙储备的库存空虚。而“饥饿激素”FGF-21所表现出来的对生殖系统和钙离子循环的抑制,似乎和我们在危险情况下逃命时机体做出的反应类似,是一种身体在食物缺乏(或者是以为自己处于食物缺乏状态)条件下的一种适应性反应。因为我们的机体许多的通讯是靠激素传递的,所以过量分泌的“饥饿激素”FGF-21可能让我们的身体(应该也是下丘脑)以为我们处于一个食物缺乏的状态,所以,它可能通过某种机制,减缓了对骨骼对钙的吸收,以保证更加重要的器官和组织不会因缺钙而失去功能,而与此同时,骨骼中的钙库存可能还在正常释放与流失,所以最终表现出来的就是实验小鼠严重的骨质疏松。所以这些小鼠的骨质疏松和孕期妇女的骨质疏松的原因是很相似的。尽管得克萨斯大学的研究者对FGF-21所表现出来的延长小鼠寿命很兴奋,但是,我对FGF-21是否真的可以让我们健康的长寿表示怀疑,我相信应该有更加深刻而且独立的机制在作用,FGF-21只是触发了这套机制的一部分而已。
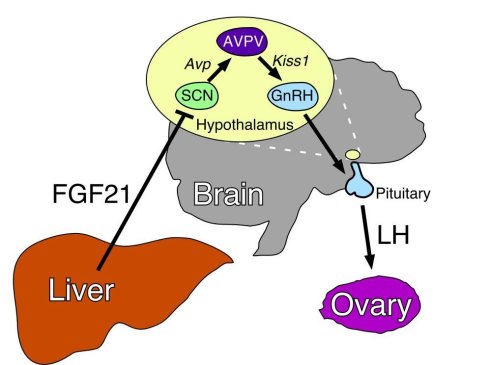 图.50 由肝脏(Liver)分泌的”饥饿激素” FGF-21 ,在雌性小鼠的内分泌系统中,信号传递是通过下丘脑(hypothalamus)——脑垂体(Pituitary)——卵巢(Ovary)生殖轴,造成雌性小鼠的性腺的衰退。
图.50 由肝脏(Liver)分泌的”饥饿激素” FGF-21 ,在雌性小鼠的内分泌系统中,信号传递是通过下丘脑(hypothalamus)——脑垂体(Pituitary)——卵巢(Ovary)生殖轴,造成雌性小鼠的性腺的衰退。同样可以对生殖系统产生抑制,并且与肝脏分泌的“饥饿激素” FGF-21类似的,是肠胃也可以分泌“饥饿激素”。Ghrelin 就是一种由主要由肠胃分泌的“饥饿激素”,胃肠,主要是胃,在空腹状态下会产生大量的这种激素。2013 年10月,伦敦帝国学院医学研究所的Alexander N. Comninos团队在《牛津期刊》旗下的human reproduction update发表了一篇文章《肠道与脂肪分泌的激素与生殖的关系》 [26],描述了生殖系统与上述两种激素的关系和作用机制。如图所示,肠胃分泌的“饥饿激素”Ghrelin 信号在雌性和雄性内分泌系统的生殖轴上,对生殖系统也是有强抑制作用的。
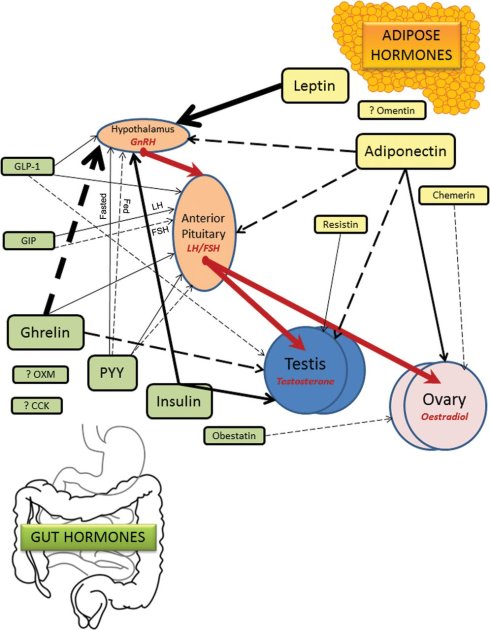
图51. 肠道分泌的激素和脂肪分泌的激素与生殖之间的关系。图中的实线代表促进作用;虚线代表抑制作用;粗线表示作用强。图中我们可以看出,肠胃的“饥饿激素”Ghrelin信号会通过下丘脑(hypothalamus)——脑垂体(Anterior Pituitary)——生殖系统睾丸(Testis)和卵巢(Ovary),对雄性和雌性的生殖系统造成强抑制。
与肝脏分泌的FGF-21类似的,肠胃产生的Ghrelin如果过量分泌的话,也可能导致骨质疏松。多数临床研究支持Ghrelin与骨量、骨密度正相关,也就是会促进骨骼生长。但是在神经性厌食症患者身上观察到的Ghrelin反常升高可引起生长激素抵抗和促肾上腺皮质激素升高,进而导致全身性骨流失以及青少年峰值骨量获得性缺失[27]。也就是说,过量分泌的Ghrelin和过量分泌的FGF-21一样,都会导致骨质疏松。
至于过量分泌的饥饿激素Ghrelin 是否能够如另一种饥饿激素FGF-21一样,也可以导致小鼠的寿命的极大延长,我没有看见相关的实验报道。相信主要还是因为这是一个需要耗时超过3年甚至4年的昂贵实验。FGF-21对小鼠的寿命延长实验是在2007年开始,2012年才出来结果。不过,在2007年左右,就已经有人开始相信Ghrelin可能可以延长寿命了。2008年11月《Life Enhancement Magazine》杂志发表了一篇文章《Ghrelin: A Mere Hormonal Appetite Stimulant or a Possible Anti-Aging Molecule》,(Ghrelin: 一个纯粹的荷尔蒙食欲兴奋剂,或者是一个可能的抗衰老分子),文中提到有证据表明,衰老可能和Ghrelin的分泌减少或者Ghrelin受到阻碍相关[28][29]。
既然空腹时产生的某些饥饿激素可以让我们长寿,那么,如果吃饱了饭后分泌的某些激素的分泌不足,会不会也让我们长寿呢?答案是肯定的。和上述两种在空腹时大量分泌的“饥饿激素”相反,我们吃饱了饭也是会大量分泌许多激素的。主要由肝脏分泌的IGF-1生长因子 (insulin-like growth factors-1)便是其中一种。IGF-1可以被译作“类胰岛素生长因子-1”,因为它的结构和功能都和胰岛素很相似,它是人体内肝细胞、肾细胞、脾细胞等十几种细胞自分泌和旁分泌的产物(也就是说人体内细胞本身就含有IGF-1)。早在1993年,加州大学的著名分子生物学家Cynthia Kenyon第一个发现了如果线虫体内IGF-1分泌不足,或者胰岛素/IGF-1受体因为基因变异而削弱(也就是线虫的daf-2基因出现突变),会使得线虫的寿命延长为普通线虫的两倍,这是非常显著的寿命的延长。这项研究也从此开启了通过寻找线虫的基因突变研究衰老机制之门。进一步的研究发现,如果胰岛素/IGF-1受体基因变异,使得受体功能削弱,那么我们可以在包括苍蝇、小鼠、甚至人类身上都观察到寿命的延长[30]。一项对世界各地的90-100岁老人的调查发现,许多人身上都发现了这个基因突变从而导致了这个受体都受到削弱。所以,体内低水平 的IGF-1或者胰岛素/IGF-1 受体受到削弱,是一个已经被大量证据证实的,可以延长寿命的因素。
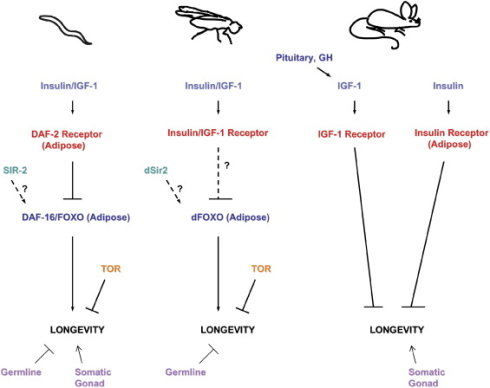
图52. 与IGF-1相关的基因和信号通道被证明影响到多种生物的寿命。图片来自: http://dx.doi.org/10.1016/j.cell.2005.02.002
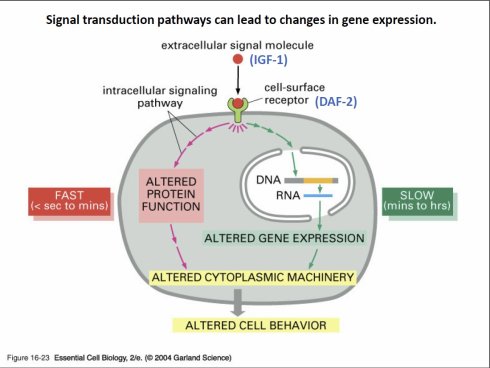 图53. IGF-1因子和细胞表面上的IGF-1受体,以及信号在细胞内的传导
图53. IGF-1因子和细胞表面上的IGF-1受体,以及信号在细胞内的传导其实IGF-1的分泌不足或者受体削弱,简单来说,所代表的含义,是和饥饿激素过量分泌是一样的。都意味着食物短缺(或者让机体以为自己处于食物短缺,当然,实际情况可能要更复杂)。而且有趣的是,在哺乳动物身上观察到,低水平的IGF-1和“饥饿激素”过量分泌一样,在延长寿命的同时,都会导致小鼠生殖发育的迟缓[31] [32],以及骨质疏松[33]。不过比饥饿激素会导致雌鼠不育好一些的是,它们的生殖系统最终还是会发育的,并不会因此不育。这其实很好理解:IGF-1水平低下代表的可能是糖和脂类等等的摄入不足,这和“饥饿激素”代表的肚子空空,完全没有食物显然轻微许多。所以,饥饿激素过量分泌会导致不育,而IGF-1水平低下只是延缓生殖,毕竟,对于生命来说,生殖才是第一要务。
有趣的是,线虫的IGF-1因素所致的寿命延长,似乎没有以牺牲健康为代价。一开始大家以为IGF-1虽然可以延长寿命,但是,它将是不健康的,比如细胞内线粒体的老化它就无法避免,所以它的老年必定行动迟缓,晚景凄凉。但最新的一则研究则颠覆了这种假设。2015年11月20日发表于《Nature Communications》的一篇文章[34],揭示了IGF-1受体受到削弱的线虫不但能够长寿,而且也更健康。这篇文章的意义在于找到了一种可以有效描述线虫活性和健康状态的方法,并对比了IGF-1受体基因突变的线虫与普通野生线虫的健康状态。他们的研究结论是,IGF-1受体基因突变(也就是线虫的daf-2基因出现突变)的线虫,更健康,更有活力,并且表明它们的细胞的线粒体也更健康。这项研究实际上揭示了线虫的生命中内含的,可以健康的延长寿命的潜力。
说一个题外话,即便是如Cynthia Kenyon 这样的大牛,也有一个我认为是错误的观点:她认为线虫获得寿命延长主要是基因突变所致,所以她曾经提出一个问题: 从线虫到人类,寿命提高了几千倍,假设线虫是我们的祖先,那么需要积累多少个基因突变才能达到这样的效果呢?我认为她这样的想法是错误的,她还没有意识到线虫们的长寿,是来自于生命内含的那种“永生”的能力,线虫要做的,只是抑制这种能力,并在必要的时候对寿命这个变量进行调整。而且大量的证据也证明了这一点:(1)饥饿所致的线虫寿命延长就不涉及到任何的基因突变,它们也可以获得如daf-2基因突变的线虫一样的两倍的寿命;有研究表明,线虫由饥饿获得的长寿特点,也可以通过Small RNA诱导的基因静默获得,并以表观遗传的方式遗传,并且可以遗传几代[35]。Daf-2基因突变所触发的长寿,其实是个“作弊”的结果,它靠削弱胰岛素/IGF-1受体的功能获得,它其实并不是一个多么稳定的结构,这可以从大部分线虫并不拥有这个基因突变可以看出,这种突变不具有演化优势。不过这个基因突变的意义在于告诉我们:我们可以通过“作弊”的方式骗过我们的死亡机制。(2)线虫的生殖细胞也是永生的,它也无需任何基因突变;(3)线虫的近亲,多次繁殖的美洲钩虫就有超过15年的寿命,线虫的祖先,水螅则可能接近永生。它们都无需演化到人这么高级,就已经可以获得和人类一样,甚至远超我们寿命的长寿了。
可以延长线虫寿命的方法其实还有许多,比如前面已经提到,直接抑制线虫的生殖系统,它们的寿命也可以获得延长。典型的例子是具有glp-1(e2141)基因突变的线虫没有生殖细胞系,它们的寿命也可以获得延长[36]。另外,美国托马斯杰斐逊大学(Thomas Jefferson University)生物化学与分子生物学系的研究人员发现线虫中单一蛋白质的水平就可以决定线虫的寿命。线虫出生时如果没有这种被称为抑制蛋白(arrestin)的蛋白质,那么它的寿命要比正常情况大约长三分之一;如果体内有三倍这种抑制蛋白的线虫,它的寿命则会缩短三分之一。[37] [38]
为什么过量分泌的FGF-21、Ghrelin等“饥饿激素”或者IGF-1水平低下,在长期作用下,抑制了生殖系统的同时,会延长寿命呢?从演化的角度上说,生殖系统被抑制的原因只是和我们在逃命时抑制生殖系统类似,主要为了缓解食物短缺的燃眉之急,以保证机体的最低正常运转。不过,生殖毕竟还是生命的最终使命之一,不完成这个使命,它的生命就没有意义。所以,它必须要调整自己的寿命和死亡机制的触发时间,同时抑制需要消耗大量能量的器官,尽可能的延长寿命,熬到食物重新丰富的那一天,以完成生殖的任务。这是生命对食物短缺这种常见问题的一个适应机制。另外,对于人类来说,饥饿与寿命的关系,可能更加复杂。我们一方面可以观察到食物的丰富,可能会导致少年儿童的性早熟;另一方面,压力的降低却又可能延长寿命。而且每一个人生活经历都是不一样的,在众多复杂因素的共同作用下,具体到某一个个体,会有什么效果,其实还很难说。所以,现在的研究发现,通过饥饿的方法,似乎并不会让人类的寿命增加多少。
小结:从上述的压力导致寿命的减少,和饥饿对寿命的延长,以及各种基因突变,或者各种激素对寿命的影响,为我们揭示了几个重要的事实:
(1)动物拥有某种潜力使其较之正常状态存活得更长久——这个事实已经受到了广泛的承认。同时也揭示了寿命长短是可调的
(2)这些寿命的延长或缩短,或多或少,都与生殖相关。
(3)有许多不相关而且相对独立的因素,都可以导致寿命的延长。这暗示,真正控制寿命长短的机制,虽然和这些因素都有关系,却是一个和它们完全独立的机制,上面提到的这些导致寿命延长或者缩短的因素,只是恰好触发了这个独立的寿命控制机制而已。
3.2.4 关于表观遗传和压力的一些争议。
关于表观遗传和压力的关系还是存在一些争议的。毕竟,表观遗传还未被证明在任何外界压力下都会产生性状改变,它不能够像DNA遗传那样稳定可靠,另外,一些缺失的环节仍然有待发现。例如有实验表明,表观遗传的印记在没有环境压力的数代之后,可能会渐渐丢失。
当许多人在诟病并强调表观遗传的不稳定和不可持续性时,有没有想过,其实很可能,不稳定或许才是表观遗传所要强调的功能和特点啊。
很可能表观遗传的目的之一就是生物在演化过程中,它需要一个可以调节的临时变量,以此适应短期的环境变化。首先在生物生活的时候,遇到压力后,它可以马上就在自己的体内改变一些相关的甲基化水平等等,以此适应一些突然的环境变化;如果变化消失,这些甲基化也可能消失并且不将其遗传。如果这个压力强度大且持续,就有选择的将它遗传(注:是否选择遗传应该还有其他的一些机制,或许会有根据历史经验选择出来的优先级等等,因为有证据表明,基因本身或许可以决定自身是否产生甲基化,也就是说,基因自身可以控制自己的表观遗传 [39])。通常生物在后天获得的表观遗传并不持久,这其实很好理解。因为假设表观遗传一旦获得,就会长期遗传,那么当环境又变回去了,它又怎么重新适应呢?所以,当环境再次变化,压力减轻后,表观遗传即便不是完全逐渐清除,也是应该被清除一部分的。简单的说,如果环境压力消失,该物种干嘛还要着急的承担巨大的机会成本,并消耗巨大能量,去缩短寿命,甚至自杀呢?毕竟,自杀和重启系统是要消耗包括机会成本在内的巨大成本的,况且周围环境是否能够提供足够它们这样折腾的食物与能量也是一个问题。 当压力消失时,也就是环境再次变化时,将已经不适应环境变化的表观遗传的印记逐渐抹去,也是适应自然选择的。
3.2.5 关于压力、生殖、遗传三者之间对寿命的影响的一个小结
关于压力、生殖、遗传三者之间对寿命的影响,我认为一个合理的演化论的解释是: 如果动物遭遇到短期的压力,它需要启动一个应急机制,让自己变得更强壮,更具有竞争力。但是,如果这样做没有效果,它继续遭遇到持续的强大压力,这就意味着他可能遇到一个非常强大的生存威胁,这或者是环境,或者是天敌等等造成的。关于压力传递途径,除了它们自己的生活经历以外,我相信有相当部分是由信息素传播的。当它们同类遭遇到危险甚至死亡的时候,释放出来的信息素可以传遍整个区域,压力也因此传播开来,虽然这种压力我们人类完全感受不到,但是对于它们的同类来说,却是一种有形有质的压力,并且压力的强度和信息素的浓度相关,也就是我们通常所谓的,一种死亡的气息。压力之下,它就可能需要适当的提高敏感度,同时加快生长发育和死亡速度,缩短寿命,加快世代交替。因为如果不加快世代交替,它很可能在没有生殖之前,也就是没有传宗接代之前,就已经被天敌或者环境消灭。这样发展下去,它的整个种群都有被灭绝的危险。所以,它加快世代交替,提前死亡是可以获利的,这样的个体也会被自然选择所选择出来。在生物面临压力的同时,在它还活着的时候,就通过DNA甲基化等等表观遗传反应,对自己进行适应性改变,以此同时,在世代交替的时候,将这些改变有选择的遗传给后代,这样,它后天采集到的信息就在后代身上完成了一个对变化了的环境的信号反馈循环。至于世代交替速度到底应该多快,自然选择会告诉它。世代交替过慢,可能会在生殖之前就被消灭,它不会有后代;但是,世代交替过快,也可能因为能量消耗过大,同时也因为机会成本的丧失,或者是食物的缺乏,而导致来不及世代交替而被淘汰。不过,好在演化是以个体为单位进行的,那些世代交替速度刚好落在速度最恰当的区间的个体,会被自然选择选择出来。
至于对饥饿压力的适应,则相反,它需要延长寿命并抑制包括生殖在内的许多机能以应对食物短缺。因为生殖本身以及生殖后的后代的生长发育都是高耗能的生理活动,在食物短缺时期,抑制生殖能力,以避免母体和后代因食物短缺而死亡,是对饥饿所造成的压力的一个正确的反应,这样的个体会被自然选择所选择出来。与此同时,表观遗传也要介入,饥饿所造成的压力,在造成母体长寿的同时,也是可以遗传的,这导致了后代的长寿,当然,这是一个临时变量,是可以修改的,所以,表观遗传通常只有几代的效果。2014年7月,美国哥伦比亚大学发表于《Cell》期刊的一项研究表明,线虫的饥饿经历,在延长自己寿命的同时,也是可以通过Small RNA诱导的表观遗传遗传给后代的,使得后代的寿命获得延长,而且这种遗传可以延续好几代。[35]
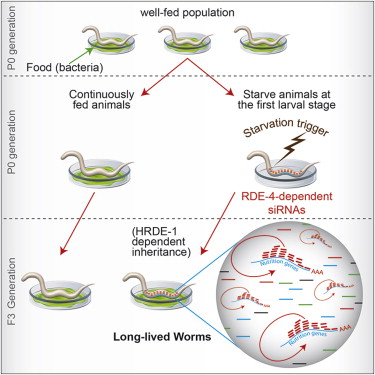 图54. 线虫由饥饿获得的长寿特点,可以通过Small RNA诱导的基因静默获得,并以表观遗传的方式遗传,而且可以遗传几代
图54. 线虫由饥饿获得的长寿特点,可以通过Small RNA诱导的基因静默获得,并以表观遗传的方式遗传,而且可以遗传几代所以,结果就是,压力、生殖、遗传(包括表观遗传)通过相互作用和相互合作,获得了一个完美的对外界变化环境的反馈循环。
当然,这所有的一切,都有一个前提,那就是,寿命长短是双向可调的,这是由生命所内含的“永生”属性所决定的。而且生物拥有一个独立的调整自己寿命的机制,它可能和许多因素都有联系,不过这套机制本身是相对独立的。上面的大量讨论和证据,其实都已经指向了这一点。并且,相信生物的寿命绝不是某个单个因素或者单个基因控制的,现在的大部分研究,可能都只是如管中窥豹一般,只看见了豹子身上的一两片斑纹,而内在的更加深刻的全局机制,也就是那一整头豹子,尚隐藏在迷雾当中..........
待续...........请点击:死亡起源(十三)
备注与参考文献
[23] Yuan Zhang,1 Yang Xie,2 Eric D Berglund,3 Katie Colbert Coate,4 Tian Teng He,5 Takeshi Katafuchi,1 Guanghua Xiao,2 Matthew J Potthoff,4 Wei Wei,1 Yihong Wan,1 Ruth T Yu,6 Ronald M Evans,6 Steven A Kliewer,7,* and David J Mangelsdorf4, The starvation hormone, fibroblast growth factor-21, extends lifespan in mice,eLife. 2012; 1: e00065. Published online 2012 Oct 15. doi: 10.7554/eLife.00065
[24] Starvation hormone markedly extends mouse life span, without need for calorie restriction. https://www.sciencedaily.com/releases/2012/10/121016103411.htm
[25] Bryn M. Owen,1 Angie L. Bookout,1,2 Xunshan Ding,3 Vicky Y. Lin,1 Stan D. Atkin,1 Laurent Gautron,2 Steven A. Kliewer,1,3,* and David J. Mangelsdorf1,4,FGF21 contributes to neuroendocrine control of female reproduction,Nat Med. 2013 Sep; 19(9): 1153–1156. Published online 2013 Aug 11. doi: 10.1038/nm.3250
[26] Comninos AN1, Jayasena CN, Dhillo WS. The relationship between gut and adipose hormones, and reproduction.,Hum Reprod Update. 2014 Mar-Apr;20(2):153-74. doi: 10.1093/humupd/dmt033. Epub 2013 Oct 30.
[27] WANG Xue-peng,HAO Yong-qiang, Ghrelin与骨质疏松相关性的研究进展,中华骨质疏松和骨矿盐疾病杂志,2010年12月第3卷第4期
[28] Life Extension News: Ghrelin: A Mere Hormonal Appetite Stimulant or a Possible Anti-Aging Molecule. http://www.life-enhancement.com/magazine/article/1980-ghrelin-a-mere-hormonal-appetite-stimulant-or-a-possible-anti-aging-molecule
[29] 6.Smith et al. Ghrelin receptor (GHS-R1A) agonists show potential as interventive agents during aging. Ann NY Acad Sci 1119:147-64 (2007).
[30] Kenyon CJ1. The genetics of ageing. Nature. 2010 Mar 25;464(7288):504-12. doi: 10.1038/nature08980.
[31] Yuan R1, Meng Q, Nautiyal J, Flurkey K, Tsaih SW, Krier R, Parker MG, Harrison DE, Paigen B.,Genetic coregulation of age of female sexual maturation and lifespan through circulating IGF1 among inbred mouse strains. Proc Natl Acad Sci U S A. 2012 May 22;109(21):8224-9. doi: 10.1073/pnas.1121113109. Epub 2012 May 7.
[32] Delayed female sexual maturity linked to longer lifespan in mice, ScienceDaily,https://www.sciencedaily.com/releases/2012/05/120507154103.htm
[33] http://baike.baidu.com/view/2040048.htm
[34] Jeong-Hoon Hahm, Sunhee Kim, Race DiLoreto, Cheng Shi, Seung-Jae V. Lee, Coleen T. Murphy & Hong Gil Nam,C. elegans maximum velocity correlates with healthspan and is maintained in worms with an insulin receptor mutation,Nature Communications 6, Article number: 8919 doi:10.1038/ncomms9919 Received 25 June 2015 Accepted 15 October 2015 Published 20 November 2015
[35] Oded Rechavi,Leah Houri-Ze’evi, Sarit Anava, Wee Siong Sho Goh, Sze Yen Kerk, Gregory J. Hannon, Oliver Hobert,Starvation-Induced Transgenerational Inheritance of Small RNAs in C. elegans,DOI: http://dx.doi.org/10.1016/j.cell.2014.06.020
[36] David Vilchez,Ianessa Morantte, Zheng Liu,Peter M. Douglas,Carsten Merkwirth,Ana P. C. Rodrigues,Gerard Manning & Andrew Dillin,RPN-6 determines C. elegans longevity under proteotoxic stress conditions Nature 489, 263–268 (13 September 2012) doi:10.1038/nature11315 Received 08 August 2011 Accepted 13 June 2012 Published online 26 August 2012
[37] sciencedaily,New protein involved in longevity identified. https://www.sciencedaily.com/releases/2010/05/100507161423.htm
[38] Palmitessa A1, Benovic JL.,Arrestin and the multi-PDZ domain-containing protein MPZ-1 interact with phosphatase and tensin homolog (PTEN) and regulate Caenorhabditis elegans longevity. J Biol Chem. 2010 May 14;285(20):15187-200. doi: 10.1074/jbc.M110.104612. Epub 2010 Mar 5.
[39] Baubec T1, Colombo DF1, Wirbelauer C1, Schmidt J1, Burger L2, Krebs AR1, Akalin A1, Schübeler D3.,Genomic profiling of DNA methyltransferases reveals a role for DNMT3B in genic methylation. Nature. 2015 Apr 9;520(7546):243-7. doi: 10.1038/nature14176. Epub 2015 Jan 21.
通宝推:lilly,铁手,唵啊吽,浣花岛主,joomla,决不倒戈,diamond,keynes,joyfm,河区分,明心灵竹,迷惑不解,陈王奋起,pattern,石狼,李根,北纬42度,家园 真是生于忧患,死于安乐阿。 有用才能永生,无用必然灭亡。
家园 不知道古中国人从哪个外星人那听来的,顺者生育,逆者成仙 其实上代的死亡,对下代的生存是有进化优势的, 自然就会进化出使生物完成生育后,提前死亡的基因。生育提供多样性,死亡消灭垄断,一代一个进程。
家园 张三丰《无根树》:顺为凡,逆为仙,只在中间颠倒颠。 FGF-21是一种所谓的“饥饿激素”,...,这些FGF-21长期过量分泌的小鼠的平均寿命要比普通小鼠增加30%,特别是对于雌鼠来说,其寿命要增加40%之多!FGF-21在导致雌鼠不育的同时,却也使得雌鼠获得超过雄鼠的寿命的延长看来闭关、辟谷、马阴藏相这些东西,有科学依据了.....

 通宝推:铁手,
通宝推:铁手,
家园 实实在在的好文章 西河好久没有这样的东西了
不得不献宝,即使通宝数已经快到0了
家园 死亡起源(十一)—— 压力与寿命 续上,死亡起源(十)
在我们对生殖对衰老和死亡的影响有了一个初步印象后,我们继续观察另外一个因素:压力,然后以压力为中心,讨论生殖、压力、遗传这三者与生物寿命的关系。
3.2.3. 压力与寿命的关系。
首先,这个“压力”指的是生物学意义上的压力(biological Stress)。现在关于生物学压力的研究已经非常丰富了,我们只要在维基百科上查“Stress (biology)”条目就可以获得大量的信息。
其次,我们需要要知道,压力和“表观遗传学”的关系,现代的表观遗传学告诉我们,许多压力所导致的变化是可以遗传的。实际上,许多表观遗传学的研究,就是靠向被研究对象施加各种压力来获得的。举一个比较新的例子,比如2015年10月,美国国家科学院院刊PNAS 刊登了宾夕法尼亚大学的研究者们的发现,父亲经受的压力会改变小鼠精子中的miRNA,进而影响其后代的大脑发育[18]。另外许多关于DNA甲基化的表观遗传实验也是通过向各种动物施加各种压力来获得的。
第三,我们现在要讨论的压力本身的含义可能要很广泛一些。这里所谓的压力,应该包含所有生物体可以感受到的信息,这包括各种感觉:冷、热、触觉、痛觉、味觉、嗅觉、听觉、视觉、饱了、饿了、湿度、光、空气、磁场、周边环境,各种相关生物(捕食者、被捕食者、竞争者等等),以及各种动、植物分泌的信息素等等所传递的信号,总之,由这些信号所造成的综合压力。
生物还有许多压力传播手段是我们人类所不能感知的,比如“信息素”就是其中一种。信息素(pheromone,音译作费洛蒙),也称做外激素,指的是由一个个体分泌到体外,被同物种的其他个体通过嗅觉器官(如副嗅球、犁鼻器)察觉,使后者表现出某种行为,情绪,心理或生理机制改变的物质。它具有通讯功能。几乎所有的动物,也包括许多植物都证明有信息素的存在。它大概可以分为警报信息素、追踪信息素、性信息素、聚集信息素、空间信息素、安慰性信息素等等。许多动物,包括昆虫和一些植物,在遇到危险或者死亡之时都会释放警报信息素。所以,在一片我们看起来祥和宁静的丛林里,那里的空气中其实是充满了只有相关物种自己才能解读的各种压力信号的。
另外,植物也可以和许多动物一样,利用“信息激素”进行通讯来传递报警和压力信号。有些植物,当动物在吃它们的时候,这些植物会分泌警戒费洛蒙使得相邻的植物产生单宁酸,而单宁酸会使得草食动物觉得植物的口感变差,变得不好吃[19]。由此可见,通过信息素,压力也可以在植物的种群中获得广泛的传播。
概括的说,这个“压力”就是生物体对所有它能够涉及和感知的,与周边环境有关的信息的处理,以及各种相关的反馈。总而言之,就是指生物所处的环境对这个生物造成的各种刺激的一个总和。如果压力是这样定义的话,那么,这个“压力”的概念,所涉及的范围之广,应该超出了人类目前的知识范围了。这个概念虽然已经超出了我们的知识范围和能力,不过,我们还是可以讨论一下简化版的压力反应的。
我们通常对于象人类这样的高等生物的压力反应指的是人们在压力面前,在大脑的控制下,交感神经和副交感神经交替作用,于此同时,大脑分泌各种脑激素,以此调控人体内分泌系统的各种激素的分泌,通知各个组织器官和细胞做出适当反应,以此达到对外界环境的某种适应性反应。
我们首先思考一下,在我们的身体中,哪个部位才是对“广义压力”的反应中枢呢?答案很明显,显然应该是我们的神经中枢——大脑。我们的大脑主要通过两套系统来与身体通讯,以此来获得对身体的各种调控。首先,作为我们的神经中枢,它可以通过我们所熟悉的神经系统对压力和各种刺激进行反映;其次,大脑作为我们内分泌系统的主腺,它也通过内分泌系统,在大脑的下丘脑(Hypothalamus)分泌各种脑激素作为信使来调控脑垂体(pituitary)等,并通过脑垂体产生各种激素,再刺激包括肾上腺(adrenal gland)在内的各内分泌腺体产生其他的激素,以激素作为“信使”来与身体各部分通讯,调控身体各部分对身体的内环境和外环境做出各种适当的反应。这个常见的压力信号通道就是所谓的下丘脑——垂体——肾上腺 (HPA) 轴(Hypothalamic-pituitary-adrenal (HPA) axis)信号通道。虽然实际过程其实非常复杂,还会涉及内分泌——神经——免疫系统三大系统的交互作用,但是简单的说,大概就是这么一个过程。
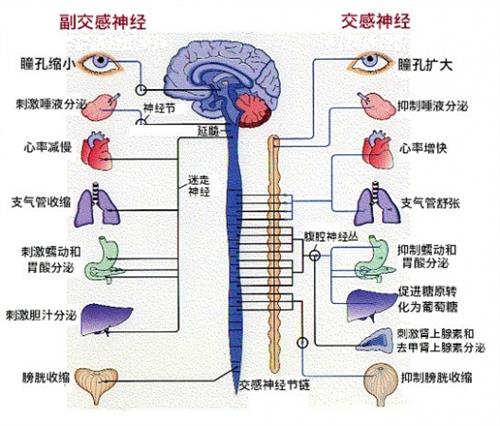
图43. 我们的交感神经与副交感神经在压力下的反应。在压力面前,在大脑的控制下,交感神经和副交感神经交替作用,对身体的不同系统,或激励或抑制。一个有趣的例子是,当一个女孩遇见一个一见钟情的对象的时候,你虽然不能直接感受到她如小鹿般乱撞的心跳,却可以从她的眼眸中看见她迅速扩大的瞳孔。这是因为,此时她体内的交感神经占据了主导,副交感神经受到抑制,与此同时,她体内的肾上腺素的水平也激升,使得她心跳加快,她可能还会感觉到嘴唇发干,那是因为唾液腺被抑制的缘故,同时她的大脑会产生大量的多巴胺,多巴胺会使她进入一种欢愉的状态..........
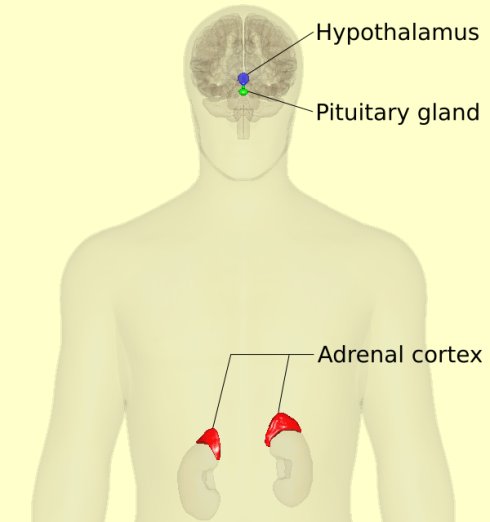 图44. 内分泌系统在压力下的反应,一个常见的压力信号通道,是通过所谓的下丘脑——垂体——肾上腺 (HPA) 轴(Hypothalamic-pituitary-adrenal (HPA) axis)传递的。
图44. 内分泌系统在压力下的反应,一个常见的压力信号通道,是通过所谓的下丘脑——垂体——肾上腺 (HPA) 轴(Hypothalamic-pituitary-adrenal (HPA) axis)传递的。进一步讨论之前,我们需要先大概了解一下什么是“激素”(荷尔蒙,hormone)。激素是指体内的某一细胞、腺体或者器官所产生的可以影响机体内其他细胞活动的化学物质。仅需很小剂量的激素便可以改变细胞的新陈代谢。简单的说,“激素”是一种可以将信号从一个细胞传递到另一个细胞的化学信使。
其实我们在讨论大肠杆菌、酵母菌以及领鞭虫这些单细胞生物的时候,就已经提到了它们的细胞间通讯了。这些细菌之间也会出现一些合作的,既然要合作,那么它们必然要互相通讯。这些细菌会分泌一些小分子化合物,释放到周边环境中,而另外的细菌身上,则有这些化合物的信号感应器,通过这些信号接收器,可以获得信号,它们以这样的方式,实现细胞间的通讯。而到了高等动物体内,大多数细胞都可以产生一种或多种分子,作为信号分子给其他的细胞传递信号,与此同时,几乎所有的细胞,根据受体数量和种类的不同,都可以有选择的接受到许多不同的相关信号。我们体内所有的细胞,其实都浸泡在一个信息的海洋当中,我们体内的细胞,每时每刻都在进行着大量的通讯。
由此,我们可以知道,我们的细胞,不管是一起协调合作,或者是要集体自杀,都是需要通讯的,而要做到这些,显然离不开这套通讯机制。
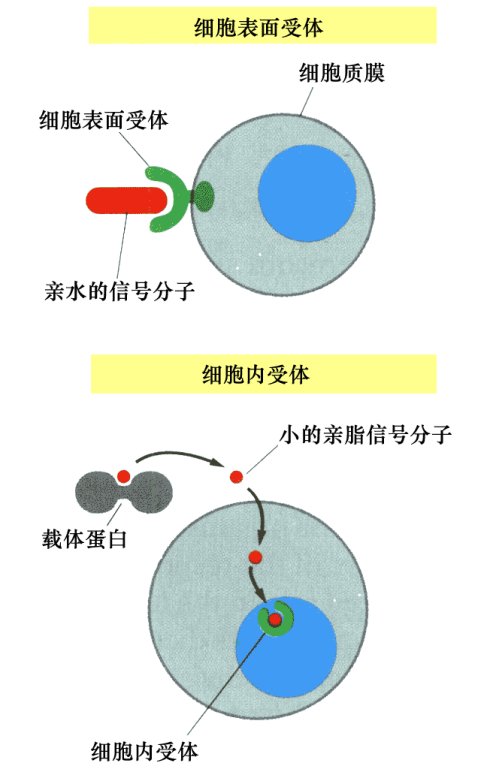 图45. 一个简化的示意图,细胞表面以及内部的受体,以及与这个受体吻合的小分子(激素)是如何吻合并传递的。细胞可以通过这种方式进行信号传递和通讯
图45. 一个简化的示意图,细胞表面以及内部的受体,以及与这个受体吻合的小分子(激素)是如何吻合并传递的。细胞可以通过这种方式进行信号传递和通讯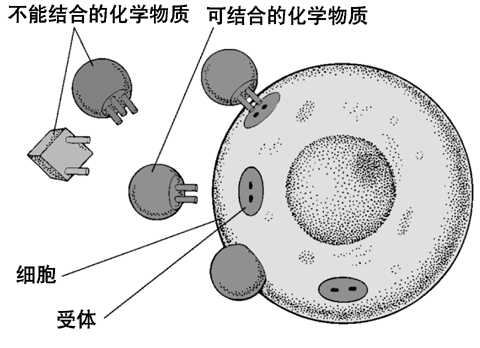 图46. 细胞表面和内部都有许多不同的受体,同时,细胞以及腺体分泌的激素也有许多种。有些细胞只有某几种受体,它只能接受某几种激素的信号,它们之间的关系,就好象钥匙与锁的关系一样。
图46. 细胞表面和内部都有许多不同的受体,同时,细胞以及腺体分泌的激素也有许多种。有些细胞只有某几种受体,它只能接受某几种激素的信号,它们之间的关系,就好象钥匙与锁的关系一样。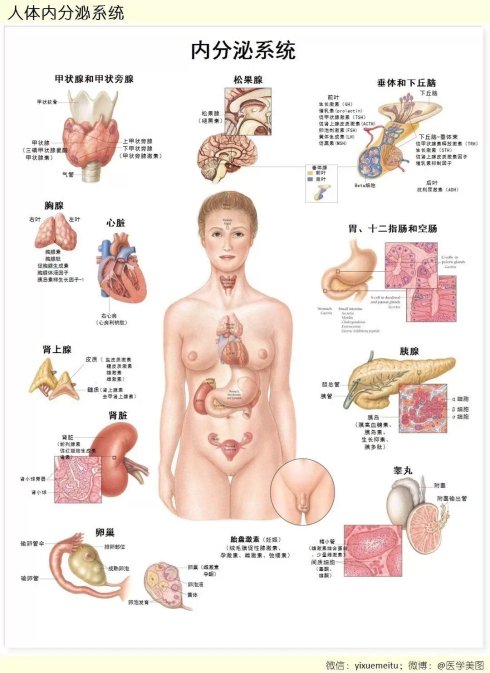 图47. 人体的内分泌系统。包括许多的器官都可以产生各种激素和化学物质。广义的内分泌系统其实范围还要更广,我们体内包括脂肪组织在内的大部分细胞都可以分泌各种小分子化合物,可以与其他组织和细胞通讯,我们的细胞是浸泡在一个信息的海洋里面的,它们每时每刻都在进行着大量的通讯
图47. 人体的内分泌系统。包括许多的器官都可以产生各种激素和化学物质。广义的内分泌系统其实范围还要更广,我们体内包括脂肪组织在内的大部分细胞都可以分泌各种小分子化合物,可以与其他组织和细胞通讯,我们的细胞是浸泡在一个信息的海洋里面的,它们每时每刻都在进行着大量的通讯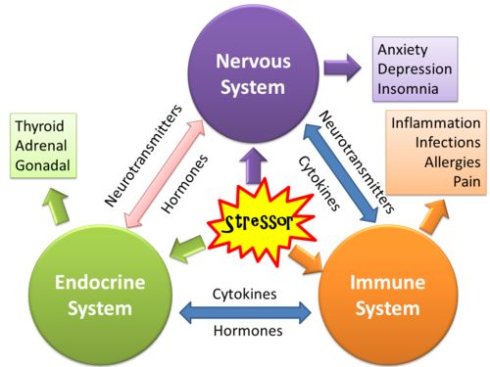 图48. 压力(Stressor) 对内分泌系统(Endocrine System)、神经系统(Nervous System)、免疫系统(Immune System)的影响,以及这三大系统相互间错综复杂的关系,它们共同构成了内分泌——神经——免疫系统网络
图48. 压力(Stressor) 对内分泌系统(Endocrine System)、神经系统(Nervous System)、免疫系统(Immune System)的影响,以及这三大系统相互间错综复杂的关系,它们共同构成了内分泌——神经——免疫系统网络所有的多细胞生物都会产生激素,植物产生的激素也被称为植物激素。动物产生的激素通常通过血液运输到体内指定位置,细胞通过其特殊的接受某种激素的“受体” (Receptor)来对激素进行反应。激素分子与受体蛋白结合后,打开了信号通路进行信号转导,并最终使细胞做出特异性反应。
激素进行信号转导包含以下几个方面1.在特定的组织中生物合成特殊的激素;2.存储并分泌激素;3.将激素运输至靶细胞;4.通过细胞膜的膜蛋白质或者胞内受体对激素进行识别;5.激素所传递的信号传递与放大:这一步最终会导致细胞的应答,而靶细胞做出反应后,产生激素的细胞可以识别出这种反应,并最终使得激素产物降解;6.激素的降解
对于哺乳动物来说,激素的作用非常广泛,它可以促进或抑制生物生长;诱发或抑制细胞凋亡;激活或抑制免疫系统;为新的生命阶段(例如青春期、更年期等)做准备;控制繁殖周期;导致情绪波动;调节新陈代谢;为交配、颤抖或逃跑等行为做准备;产生饥饿和渴的感觉;产生性兴奋或性冲动等等。由激素的各种作用可以看出,激素一定也和我们要讨论的衰老与死亡的话题相关。并且因为激素的分泌,许多都是和大脑所受到的压力相关的,由此我们知道,许多激素的分泌也是和压力相关的。
和人类一样,许多生物对“广义压力”的反应也是通过调整各种激素的分泌来实施的。
大概在1930年代,研究者就观察到一个现象:长期压力下的小鼠可以观察到免疫系统萎缩,肾上腺肥大,胃溃疡等症状。后来人们在灵长类和人类身上也观察到了这些现象。进一步的研究表明,这些现象和我们体内的“压力激素”水平变化相关。 时间又过去的许多年,现在我们已经开始知道,压力过大会伤害人体神经系统、骨骼肌系统、呼吸系统、心血管系统、内分泌系统、胃肠道系统、生殖系统等7个系统的健康。而这些现象,很大程度上都和我们的内分泌系统产生的各种激素,特别是“压力激素”相关。
在我们体内,位于肾脏上端,两个拇指大小的肾上腺分泌着三种关键激素:糖皮质激素、脱氢表雄酮(DHEA)及肾上腺素。这三种激素被称为“压力激素”。其中糖皮质激素及肾上腺素是“加压激素”,而DHEA则是“减压激素”。
其实与压力相关的激素非常多,我们仅以最研究得最深入的,也最典型的糖皮质激素来讨论。而且,在前面的讨论我们已经知道了,杀死Sockeye三文鱼和阔脚袋齁的主要杀手,也正是糖皮质激素。
糖皮质激素(gc)是由肾上腺皮质束状带细胞分泌的一类甾体激素,具有广泛的抗炎、免疫抑制和抗肿瘤活性,包括诱导T细胞和B细胞凋亡的能力。低水平糖皮质激素是维持胸腺功能的必备物质,刺激各种成熟淋巴细胞活性,加速未成熟前淋巴细胞发育为效应淋巴细胞,增强胸腺分泌胸腺激素。但是,值得注意的是糖皮质激素诱导胸腺细胞死亡模型是研究凋亡经典模型,对胸腺细胞凋亡的分子机制的认识多来源于此。由糖皮质激素诱导的细胞凋亡程序既有细胞特异性,也有共同特征,它可以快速令小鼠胸腺显著萎缩,并且使胸腺细胞减少80%以上。研究还发现生理糖皮质激素水平引起大规模胸腺细胞凋亡可以通过肾上腺切除逆转。随着程序化细胞死亡机制研究的深入,糖皮质激素这种作用机制越来越明确,因为它可以诱导DNA降解成180 bp及其倍数的dna片断,这种DNA降解规律是凋亡的典型特征。[62]
关于压力对我们身体的激素分泌和免疫系统的影响,一直有一个在演化上难以解释的现象:我们在受到短期压力的情况下,我们的大脑会控制和调节内分泌系统,导致体内的糖皮质激素水平迅速升高。短期的糖皮质激素水平升高是可以激活并加强我们的免疫系统的,这个很好解释。但是如果长期的处于压力之中,体内增加的糖皮质激素则会开始抑制,甚至摧毁我们的免疫系统。糖皮质激素甚至可以直接杀死我们体内的淋巴细胞(一种免疫细胞)。糖皮质激素可以诱导并引起大规模胸腺细胞凋亡。而且,糖皮质激素对我们的杀伤并不止这些,长期的高浓度的糖皮质激素它还会杀伤我们的脑神经细胞,损害大脑的海马体等等。所以,压力会影响内分泌系统,并且长期的压力下,我们的内分泌系统甚至会抑制并摧毁我们的免疫系统。在极端情况下,体内高浓度的糖皮质激素甚至可以直接杀死Sockyey三文鱼。
这是一件很奇怪的事情,为什么我们会演化出这么一套奇怪的机制,这是用一般的理论无法解释的。我们很难理解,为什么好端端的要将我们的免疫系统摧毁掉?对于个体的生存来说,这显然不是一个好的选择。不过,如果把这个问题放到本文一直在讨论的话题——衰老与死亡的产生来看,我们就有一个合乎逻辑的解释了。
一个合理的演化论的解释就是:生物在演化过程中,需要具备对压力的反馈能力,并通过不断调整,维持一种动态平衡。而且这种能力还必须是可以遗传的,因此遗传是实现压力反馈的一个重要环节。如果它们不具备对压力的适当的遗传反馈能力,这个物种很容易被自然选择所淘汰。而表观遗传在这种适应反馈中起到了非常重要的作用。长期的压力对于所处其中的生命来说,往往意味着一种长期的,经常剧烈变化的,需要不断适应的竞争环境。在长期的压力面前,如果不能适应,物种面临的是可能威胁整个物种延续的生存危机。所以,它们需要一方面可能通过后天修改自己的某些甲基化水平等等,修改自己的各种表观遗传来获得某种适应,同时在必要的情况下,还需要加快自己的演化速度来适应这种变化。为此它们可能需要加快自己的生命周期,加快自己的生长发育和新陈代谢,加快自己的衰老和死亡速度,通过不断的快速调节表观遗传的各种开关,不断的更新换代来适应各种变化。虽然这种加速是需要消耗能量并要付出相当代价的,不过,在自然选择这只看不见的手的微妙调节下,它还是可以找到一个符合自己利益最大化的,适合当前竞争环境的平衡点。因此,在长期的压力之下,我们的身体主动摧毁自己的包括免疫系统在内的各个系统来加快自己的衰老和死亡速度,就有了合乎逻辑的演化论上的解释了。
而反之,当一个物种处在一个相对优势的,“广义压力”较低的环境时,它可能就会选择延长寿命(包括延长生殖寿命)的策略。我们从许多大型乔木和其他生物的策略上可以知道,延长寿命在竞争中也是可以获得许多好处的。而且,延长寿命对能量的消耗也相对小许多。所以我们可以看到许多哺乳动物也可能拥有很长的寿命,比如我们人类、大象、以及寿命可能长达200年的弓头鲸(Bowhead whale),另外,通过考古发现,一些大型恐龙的寿命也很长,这可以通过分析它们的骨骼的年轮得知。这些在生态链处于一个优势地位,而且食物相对简单,不需要通过剧烈的捕猎来获得食物的动物,往往可以观察到它们会倾向于选择长寿的策略。相应的,它们的新陈代谢速度或许也会因此比小动物减缓一些。而同样处于生物链顶端的狮子是通过剧烈的捕猎来生存的,剧烈的捕猎则意味着竞争和压力,狮子甚至可能在捕猎中受重伤,所以狮子的寿命并不长,大约只有10到14年。与狮子相反的是,大象的食物是植物,弓头鲸的食物是磷虾和浮游生物,它们都没有捕猎的压力,而它们巨大的体型也保证了成年后的它们自身被天敌捕猎的压力也相对较轻。另外对于狮子来说,如果生活在动物园里面,没有压力的话,有记载其寿命可能长达34年。
对于压力来说,人类的情况要复杂许多。不过纵观人类历史,我们的总体寿命是一直在延长的,衰老速度也在变慢,相信这不仅仅是因为科技的发达以及食物的丰富,也和现代普通人的总体生存压力相对古代的普通人要低许多所致。另外,人类和大部分动物相比,已经脱离了狂野森林中,弱肉强食的从里法则了,所以人类所面临的压力,是远比绝大多数动物小的。大部分动物,不论是捕猎在还是被捕猎者,长期面临的,往往都是生死攸关的竞争和压力。再加上人类拥有动物界中最发达的大脑,人类也因此具有了哺乳动物中,非常长的,包括生殖寿命和老年寿命在内的长寿命(注: 关于哺乳动物的寿命与大脑的关系我们会在后文讨论)。当然,我们也需要意识到,演化的基本单位是个体,每一个生物个体实际都在走自己独立的演化之路。如果我们面临长期的压力的话,我们上面的讨论以及各种对内分泌系统的研究告诉我们,我们的机体或许就会悄悄提前启动那套自杀机制,导致我们的早衰甚至死亡。
与蜜蜂和白蚁的生殖与激素可以双向调节寿命的长短类似,压力对寿命的影响也是双向的。因为寿命本身就是一个可以按需要而进行双向调整的变量。生物在生命当中可能面临的诸多压力当中,饥饿也是一种很常见的压力。 不过与上述压力的作用相反的是,饥饿所造成压力却可以延长寿命。大概该上世纪30年代开始,研究者就发现,适当的饥饿可以延长生物的寿命。康奈尔大学在1934年的一个著名实验中,发现在实验室中,只要给老鼠喂低卡路里的食物,但又同时保证他们可以获得足够的营养以避免营养不良,那么,这些老鼠的寿命就可以达到预期寿命的两倍。
在过去的80多年间,许多研究机构做了大量的动物节食实验。他们发现剥夺营养可以延长包括酵母菌、蠕虫、果蝇、蜘蛛、鱼、大鼠、小鼠直到灵长类的猴子的寿命,它们的寿命可能比它们自由进食的参照组高30%到200% [21] 。
2014年6月,美国杜克大学(Duke University)的研究者们发现:如果拿走线虫(C.elegans)的食物,就会触发线虫进入一个发育停滞的状态:它们还会继续蠕动,继续寻找食物,不过它们的细胞和器官却会进入一个不会衰老(ageless)的悬停静止态。当重新恢复供应足够的食物之后,它还能继续正常发育,不过却可以因此获益,获得最高长达延长至两倍的寿命。这个研究成果于2014年6 月19日被发表在遗传学杂志《PLOS Genetics》上 [22] [21] 。
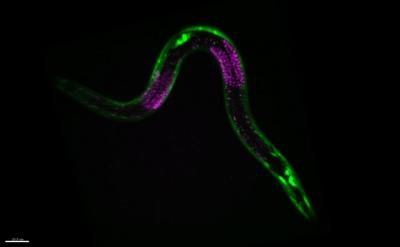
图49. 图示的是细胞被荧光染色后的秀丽隐杆线虫(C. elegans)。绿色的是肌肉细胞,红色的是生殖细胞。它们在饥饿的条件,这些细胞的发育会在一个发育节点(checkpoint)前暂停,并延缓它们的衰老。
我对于该实验中展现出的线虫可以自由调节寿命其实并不感到很特别,因为经过这么久的讨论,我们应该知道,这本是天经地义的事情,对于生物来说,寿命本就是可调的一个变量。我感兴趣的是,上述实验在无意中展示的,生殖的延迟对寿命的影响。研究员Sherwood和他同事们一开始把研究的目光集中在了线虫的幼虫发育的最后两个阶段——L3 和 L4,在这两个阶段,线虫的一些关键的组织和器官还在生长发育当中。在这两个阶段当中,线虫的生殖孔(Vulva,也就是它的生殖器)会从一个只有3个细胞的小不点成长到一个有22个细胞的小球。他们发现,当他们在线虫发育的这两个阶段中如果取走食物,线虫的生殖孔的发育就会停止在要么3个细胞的状态或者要么22个细胞的状态,(而不会是中间的,比如10个细胞的态)。随着进一步的研究,他们在线虫全身所有的组织和器官中都观察到了同样的现象。这篇文章的意义在于,发现了线虫生命的发育节点或者“检查点”(checkpoints)。它告诉我们,生命的发育不是连续的,而是有许多节点的。线虫的发育过程中有许多的“检查点”,它的机体会评估每一个检查点,看看条件是否合适,如果合适,就继续发育,如果出现如上述的食物短缺,营养不足的情况,它们的发育就会在这个检查点前面停止下来,等待条件合适再继续。而且我们也可以从这个实验看出,杜克大学的研究者所观察到的线虫的寿命延长,是已知的绝大多数的衰老和死亡机制所不能解释的。
不过,正如本文2.4中所提到的,在杜克大学这份报告发表一年后,美国西北大学的研究团队则在2015 年7月发现了线虫(C.elegans)的衰老死亡开关了[11],并找到了相关的基因,他们发现线虫的死亡不是所谓的自然衰老所致,它们的死亡是受生殖干细胞触发的开关控制的。 而且线虫的死亡开关的打开的“检查点”(checkpoint) 是在生殖系统成熟8个小时之后启动。所以,饥饿本身并不是造成线虫寿命延长的根本原因,它之所以寿命获得延长的一个可能的原因,是因为它们的发育还没有走到由生殖系统成熟所触发的死亡开关的那个“检查点”,正常情况下,没有走到那个被生殖细胞触发的死亡开关的节点之前,如果它还没有被饿死的话,或者没有被它的死亡机制的另一个触发点触发的话,线虫有相当大的可能是不会主动触发死亡开关的。也就是说,线虫的饥饿所引发的寿命的延长,只是为了可以让它可以积攒足够的能量,让它的发育可以走到生殖那个点,以完成繁殖的使命。由此我们也可以看到线虫的寿命与生殖的关系。虽然线虫的寿命与生殖相关,但是,线虫的死亡机制是与生殖独立的。而这里面更加深刻的内在原因依旧是,寿命本就是可调的,所以它才能自如调整,如果寿命是固定且不可调的话,它是不可能获得这样达到两倍的大范围的调整的。(注:线虫在切除生殖系统后,也是会死亡的,并不会因此永生。另外,有glp-1(e2141)突变的线虫没有生殖细胞系,它们虽然可以获得寿命的延长,但是也是会死亡的。这说明线虫的死亡机制虽然可以由生殖系统触发,却是一个与生殖完全独立的机制)
上述现象在演化论上的解释也很简单。生命存在的意义是什么?当然是为了活着,为了发育,为了繁殖。如果因为食物短缺,而使得自己不能获得足够的能量去发育和繁殖,也使得后代不能获得足够的能量去发育与繁殖,那怎么办?马上因为压力而自杀吗? 显然不是。生命反而要迎着这个压力去延长自己的寿命,尽量等到食物重新丰富、累积的能量足以繁殖的那一天。这也是本文一开始举例的北极灯蛾的生存策略,这是一种很正常的演化结果。
比线虫更复杂的,是哺乳动物在饥饿压力下的反应............
待续.............请点击:死亡起源(十二)
备注与参考文献
[11] Simple Flip of Genetic Switch Determines Longevity in Animals. http://www.northwestern.edu/newscenter/stories/2015/07/genetic-switch-determines-longevity-in-animals.html
[18] Ali B. Rodgers, Christopher P. Morgan, N. Adrian Leu, and Tracy L. Bale ,Transgenerational epigenetic programming via sperm microRNA recapitulates effects of paternal stress,PNAS doi: 10.1073/pnas.1508347112 November 3, 2015
[19] J.du P. Bothma, Game ranch management, fourth edition, Van Schaik publishers, 2002
[21] Strict diet suspends development, doubles lifespan of worms, http://www.eurekalert.org/pub_releases/2014-06/du-sds061314.php
[22] Adam J. Schindler, L. Ryan Baugh,David R. Sherwood,Identification of Late Larval Stage Developmental Checkpoints in Caenorhabditis elegans Regulated by Insulin/IGF and Steroid Hormone Signaling Pathways,PLOS GENETICS, Published: June 19, 2014?DOI: 10.1371/journal.pgen.1004426
[62] 钱知勉(综述),殷彦君,激素对胸腺影响的研究进展,滨州医学院学报2009年2月第32卷第1期
通宝推:lilly,铁手,唵啊吽,浣花岛主,坐言起行,河区分,李根,keynes,石狼,北纬42度,乔治·奥威尔,陈王奋起,家园 这个“糖皮质激素”和器官移植异体排斥反应有没有关系? “糖皮质激素”增加短期内增强免疫功能,长期则可能摧毁免疫系统。这个现象,从纯逻辑的角度来看,应该是“糖皮质激素”提高了免疫系统的敏感性。结果是,虽然对外界威胁反应加强了,但是对自身细胞也有了攻击。印象中白癜风和红斑狼疮都是免疫系统对自身细胞攻击的结果。
家园 有趣,那么完成生殖使命后的寿命是如何调节的呢 另外,像人这样的生殖期很长的动物,演化在对待寿命与生殖任务完成与否是怎样的一种关系。
比如听到过一种说法,人老年的各种疾病包括癌症,都是因为人的生殖任务完成之后,机体(这里有你提到的死亡基因开关的作用)触发的,因此早早晚晚都要是一身的病。但人的生殖功能可以一直延续到五六十岁,有的男人甚至更久,还是说在大型哺乳动物身上,这个死亡与生殖的关系不那么密切了。
家园 提点意见 究竟是生命存在的意义是为了永生
还是迄今为止 生命一直在追寻永生
这里的却别在于如果是后者 生命能够达到永生(或者足够长,长到不想活了) 那么是不是会追寻点别的 比如活的更好更精彩
- 复 提点意见
家园 生命的意义应该是自然环境的一部分 物理反应,能量转换,问生命意义的时候就免不了要问宇宙存在的意义,这个就目前无解了。
细菌,昆虫,屎壳郎这种生命存在的意义可以从实用视角得到解释,甚至大型哺乳动物也可以从食物链的存在得到解释,估计人要问这个问题终归还是因为人的大脑。
生命的基因观认为生命的意义在于基因的传播或者基因的永生,而基因因为只是信息,于是生命又不过是信息传播的一种方式。
不过人可能真正想问的不是生命存在的意义,而是产生大脑这种智能的意义。毕竟生命司空见惯,地球上随处可见。但是人类大脑智能这个现象,为什么产生,存在的意义是什么,想想的确让人毛骨悚然,难怪人类自古产生那么多宗教,可怜的古人逻辑无法自洽啊,现代人也很难。
家园 死亡起源(十)—— 生殖与寿命 续上, 死亡起源(九)
(注:本章涉及到一些关于“激素”和“细胞间通讯”以及细胞的自杀——“凋亡”的概念,如果不清楚,请参考下两篇:死亡起源(十一),和死亡起源(十三)因为这些概念是混在一起的,很难剥离,这篇我只是试图单独的描述生殖与死亡的关系。所以没有做过多的解释和介绍。)
简单讨论完表观遗传后,我们继续讨论我们的主要话题:衰老与死亡。前文我们已经提到了,尽管已经有了许多的研究告诉我们,我们的衰老和死亡和许多因素都有关系,而且这些因素都有大量的研究做依据,不过,穿过重重迷雾与噪声,从演化的角度上看,我观察到的与衰老与死亡相关的最重要的钥匙却是如下几项:
1. 生殖;2. 压力;3. 遗传与变异 (包括表观遗传)。
其实这三者之间的关系是相互影响的,并不能完全剥离,我现在试图把它们一个个分开来讲。
3.2.2 生殖与寿命的关系。
经过前几章漫长的讨论,我们已经观察到了一些可以揭示生命本质的有趣现象。现在,我们讨论生殖与寿命的关系。其实,在前面的讨论中,我们已经清楚的看见了生殖与寿命的关系了。比如美国西北大学的线虫衰老死亡报告,揭示了线虫的衰老和死亡,是由线虫生殖干细胞触发了一个死亡开关所致。Sockeye三文鱼的产卵后的自杀,显然也是和生殖有关。进一步讨论生殖对寿命的影响之前,我们先简单介绍一下关于生殖的一些基本概念。
生殖按分类主要可以分为两大类: 1. 无性生殖;2.有性生殖。关于无性生殖以及有性生殖的起源以及对死亡产生的影响,前面已经讨论了很多了。现在我们再进一步讨论生物的“生殖策略”。
采用有性生殖的物种的生殖策略主要又可以分为两大类,单次繁殖和多次繁殖。:
单次繁殖的生物在一生中只进行一次生殖,例如一年生植物(包括所有的谷物作物)、一些种类的鲑鱼、蜘蛛和竹子等。这些生物通常在繁殖后很快便会死亡。
多次繁殖的生物会按照连续的周期(例如每年或每季度)产生后代,例如多年生植物等。多次繁殖动物可以适应多个季节以及周期性的环境变化。
现在我们比较一下单次繁殖和多次繁殖这两种策略。理论上说,单次繁殖的策略,它们的演化速度相对会更快一些,而多次繁殖的策略,则单位能量消耗要相对小一些,机会损失也小一些。这两种策略各有优缺,至于谁优孰劣,还要放到各自具体的竞争环境中才知道。
当我们观察单次繁殖动物,比如大部分昆虫类,我们往往会发现,它们中的许多,似乎没有严格意义上的衰老现象。它们的存在的唯一目的似乎就是为了生殖,生殖一完成,它们的使命包括生命也就该结束了。许多昆虫似乎刚才还是生龙活虎,完全没有衰老迹象,而生殖完成后,往往在极短的时间内就死亡了,他们似乎也和线虫一样,不存在衰老的概念,它们的死亡只是一个开关的打开和闭合。有些单次繁殖的生物,比如许多龙舌兰属(Agave)的植物,它们的寿命可能很长,可能需要100年才能成熟。不过,它们一旦成熟并繁殖后,就会立刻死亡。单次繁殖的竹子也是类似,有一年它们开花繁殖导致的大量死亡,还唱红了一首歌:《熊猫咪咪》。至于单次繁殖的Sockeye 三文鱼,它们实际上不是严格意义上的单次繁殖动物,而是多次繁殖动物采用了单次繁殖的策略。
即便是多次繁殖的生物,(高等的哺乳动物除外),它们的存在似乎也只是为了生殖,许多多次繁殖的低等动物一旦停止生殖,往往就意味着死亡。另外,在低等的多次繁殖的生物身上,衰老现象同样也不是那么明显。前面已经讨论过了,我们可以观察得到单次繁殖的秀丽隐杆线虫只有两三周的寿命并且在生殖后快速死亡。而它们的近亲,多次繁殖的美洲钩线虫的寿命却可以长达15年,美洲钩虫日均产卵可以高达5000~10000个。我们还可以观察得到同样低等的多次繁殖的血吸虫竟具有长达几十年的寿命,这几乎和它的宿主,早期人类的寿命相当了 。而且,具有超强再生功能的血吸虫也似乎也不会衰老。另外血吸虫在中间宿主钉螺体内是进行再生性质的无性繁殖的,在终宿主体内才进行有性生殖。另外,而秀丽隐杆线虫如果进入了所谓的“dauer幼虫”状态,它也会停止老化。总之,生命之奇妙,经常让我们难以想象。
而真正意义上的衰老现象,我们可能要在哺乳动物身上寻找,在哺乳动物身上,我们可以观察得到较为严格意义上的衰老现象——就是我们所熟悉的,那种渐进性的衰老。关于这个话题我还会在文章后面讨论。
总之,我们可以观察到,生殖是生命的一个非常重要的环节,许多生物的寿命,都和生殖相关。它们为了生殖,或者如单次繁殖的线虫般,产卵后突然自杀;或者如许多多次繁殖生物一般,尽量延缓开关的打开,延长自己的寿命。有些寄生虫的寿命策略和许多大型乔木的策略有些类似,因为延长寿命符合利益最大化,所以它们就延长寿命 (推迟打开死亡开关)。所以,我们可以观察到,同属于线虫纲的秀丽隐杆线虫只有20天的寿命,而美洲钩线虫则可以有长达15年的寿命。
如果上面的例子还不足够说明问题的话,我们再观察一些例子。相信有人或许会质疑,这些生物寿命差别巨大,很可能是因为它毕竟是不同的物种,或许它们各自有各自的非常特别的基因,才导致了这么巨大的差别。那么,下面我们要观察的的例子,则会告诉大家,即便是同一个物种,甚至可能是同一个DNA拷贝,寿命也可以有巨大差别,而且这个差别,可以高达几十到100倍!
关于生殖对寿命的影响,一个有趣的例子就是蚂蚁和蜜蜂。和大多数单次繁殖的昆虫不一样,蜜蜂和蚂蚁都是“多次”繁殖的。虽然它们大多一生只受精一次,但是,它们却可以把精子或者受精卵泡存储起来,慢慢的使用,达到“多次”繁殖的目的。(白蚁或许会多次交尾,成为正真意义上的多次繁殖昆虫)。同是雌蚁的蚁后与工蚁的基因完全一样,但是一般蚂蚁的蚁后的寿命可能长达15年,是工蚁的100倍。而白蚁的蚁后的寿命甚至可能长达50到70年。蜜蜂的寿命也是类似,同是雌蜂的蜂后和工蜂之间也存在着巨大的寿命差别,蜂后的寿命可以长达4到5年,而工蜂的平均寿命只有45天。我们已经知道,蜂后和工蜂唯一的区别就是它们的在成长时所被喂养的食物不同。蜂后被喂养的是可以促进保幼激素分泌的蜂王浆,工蜂则是普通的花蜜和花粉。至于蚂蚁,我们现在已知的是蚂蚁的蚁后通过信息素抑制了工蚁的产卵能力。另外,对于蚁后自身,若是它体内存储的精子如果耗尽的话,本来似乎不会衰老,没有显露出一点衰老迹象的蚁后往往会立刻死亡。蚁后死亡以后,失去抑制的蚁群会自动由普通工蚁当中产生新的蚁后。至于雄蚁和雄蜂,它们存在的唯一目的似乎就是交配,所以它们往往会在交配后立即死亡。从蜜蜂和蚂蚁的例子我们可以看出,主流的许多解释衰老的原因或假说,似乎都无法解释蚁后与工蚁,蜂后和工蜂之间的巨大寿命差异。我们倒是可以从中清楚的观察到,生殖以及相关激素和信息素对它们寿命的巨大影响。
当然,我们这里需要再次强调的是:蜂后和蚁后之所以有这么长的寿命,内在的根本原因,并非是激素,也不是生殖,而是它本就有长寿的能力。激素其实只是一个信使,它的作用只是传递出一个信息,这个信使通知它们打开或者关闭死亡的开关。
 图41. 白蚁的蚁后、兵蚁和工蚁的尺寸对比,白蚁的蚁后的寿命最高可以达到50年以上。它们的例子告诉我们,同一个物种,甚至是同一个DNA拷贝,也可以获得大范围的寿命的调整,这个“大范围”的意思是:几十到100倍
图41. 白蚁的蚁后、兵蚁和工蚁的尺寸对比,白蚁的蚁后的寿命最高可以达到50年以上。它们的例子告诉我们,同一个物种,甚至是同一个DNA拷贝,也可以获得大范围的寿命的调整,这个“大范围”的意思是:几十到100倍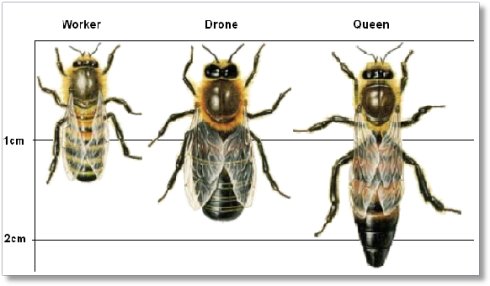 图42. 蜂的工蜂、雄峰、蜂后的尺寸对比,蜜蜂的蜂后寿命可以长达5年,相较之下,工蜂的平均寿命只有45天。蜜蜂的例子也告诉我们,同一个物种,甚至是同一个DNA拷贝,也是可以获得大范围的寿命的调整的,这个“大范围”的意思是:大约40倍
图42. 蜂的工蜂、雄峰、蜂后的尺寸对比,蜜蜂的蜂后寿命可以长达5年,相较之下,工蜂的平均寿命只有45天。蜜蜂的例子也告诉我们,同一个物种,甚至是同一个DNA拷贝,也是可以获得大范围的寿命的调整的,这个“大范围”的意思是:大约40倍关于蜂后身上发生的一些细节,我们可以从2011年4月《Nature》杂志的一篇文章[20] 中找到答案。对于蜂后来说,它是通过摄取了蜂王浆中的一种叫Royalactin 的蛋白质,而这种蛋白质最终通过一系列过程,会提高它体内的保幼激素浓度,由此引发了它的卵巢发育,身体变大等等一系列变化。做一个形象的比喻,这也就是说,在蜂后的体内,保幼激素等等激素作为一个信使,通知了它自己体内的细胞:现在大家不要急着自杀了, 我们荣幸的接到通知,委任我们为本蜂巢的蜂后,负责承担整个蜂群的生殖任务,而这需要我们延长我们这个机体的寿命。于是它们的系统就重新调整机体以及细胞的寿命,使得它整个机体的寿命获得延长。与此同时,蜂后的大颚腺还可以分泌一种信息素“蜂后费洛蒙”(royal pheromone),以此来对外抑制(准确的说是通知)蜂群里面其他工蜂的卵巢不要发育。另外,蜜蜂的幼虫和蛹也可以分泌类似的信息素来实现对工蜂的抑制或者通知。而与蜂后相反的是,白蚁的蚁后体内的机制,却可能是默认将身体调得很长寿的。当然,这需要在没有压力,包括充足的食物、工蚁的良好照顾、安全的环境等等条件下才行,离开蚁巢单独生存的蚂蚁一般只能活几天。在外激素的使用上,蚁后主要也是通过分泌费洛蒙出去,让这些激素到身体外面去抑制工蚁的长寿的能力。蚁后通过分泌费洛蒙,让这个信使进入工蚁的体内,去通知工蚁体内的细胞,让它们抑制自己的生殖能力,或许还要引发它们在适当的时主动自杀。白蚁的费洛蒙对蚁群的作用,实际上还要更复杂一些,它对白蚁的蚁后、雄蚁、兵蚁和工蚁的阶级分化和数量调节也起到了至关重要的作用。
关于激素和信息素(费洛蒙)对生命的影响,我们将会在“压力对寿命的影响”中进行比较详细的讨论。
由此,我们只要通过比较蜂后和蚁后的内激素和外激素的不同作用,就可以看出,在它们的生殖过程中,激素作为一个信使,在生物寿命的控制与反馈中起到的双向作用——它既可以延长寿命,也可以缩短寿命。而且我们也可以看出,生物体内的内在的,与激素和生殖既相关,又独立的,那种可以大范围调节自己寿命的能力。
待续........ 请点击:死亡起源(十一)
备注与参考文献
[20] Kamakura M1.,Royalactin induces queen differentiation in honeybees. Nature. 2011 May 26;473(7348):478-83. doi: 10.1038/nature10093. Epub 2011 Apr 24.
通宝推:lilly,铁手,唵啊吽,浣花岛主,河区分,北纬42度,keynes,石狼,