主题:233-Alison Woollard:生物学的七场基因革命 -- 万年看客
- 共: 💬 2 🌺 18
https://www.youtube.com/watch?v=gAOPxpm_3EA
一切科学学科都试图建立大一统理论。物理学用热动力学统合了力学与热学;爱因斯坦的相对论加强了物理大一统的前景,指出能量和物质只是同一枚硬币的两边;波粒二相性与共同参考系让我们在量子力学的背景下理解了狭义相对论。但是足以解释万物的大一统理论也就是量子引力理论依然尚未现身。相比之下,生物学已经有了自己的大一统理论,也就是达尔文在1859年发布了基于自然选择的进化理论,再加上几年后遗传学先驱孟德尔提出的遗传理论。这些遗传理论就是我们今天讲座的主题。接下来我们将会看到,这套理论基于一系列革命性的理念,由许多世界所知的最伟大科学家提出。这是个令人兴奋的故事,因为它讲述了将我们对于生命的理解提升到分子层次的革命。而且此时此刻讲述这个故事尤其令人兴奋,因为我们正在进入一个新时代。如今我们不仅理解了基因对于生命的影响,而且还在培养足以干预自身基因命运的能力。因此如今我们尤其应当理解革命的来龙去脉,革命意味着什么,我们应当如何前进。不仅只有科学家需要如此,整个社会都应当具备这方面的认识。
达尔文的理论认为自然环境下的生物差异源自生存斗争,只有最适应环境的物种才能生存。进化基本意味着随着时间流逝而发生的渐变,自然选择是进化的机制。有些变化得到了自然选择的青睐,使得生物更适合在特定环境生存繁殖,诞生更多具有同样特点的后代,比如加拉帕戈斯群岛上的达尔文雀。所以一种生物假以时日完全可以变成另外一种截然不同的生物。通过阐述基于自然选择的进化论,达尔文意识到地球上的所有生物全都相互关联,有着共同的起源。因此他构想了生命之树的理念,正如画面所示,这是达尔文亲笔手绘的生命之树。达尔文的树造型很简单,如今的生命之树则要复杂得多。当然,达尔文是正确的。所有物种确实相互联系,它们的共同祖先是四十亿年前的简单生命形态,自然选择将这些简单生命送上了各种不同的道路,由此造就了非凡的生物多样性——整整九百万种物种,这还只是曾经生存过的所有物种的一小部分。达尔文激情四射地宣称他眼中的生命“无比壮美”,充满了“无穷无尽最美丽最神奇的形态”。达尔文的自然选择理论作为生物学的大一统理论如此成功,以至于进化生物学家西奥多修斯.多勃赞斯基(Theodosius Dobzhansky)曾经说过:“若非蒙进化之光,生物学毫无意义。”
但是达尔文的理论有一个问题:达尔文无法提出可靠的机制来解释遗传,因此他的逻辑不完整。他的大一统理论在这方面失败了。达文知道生物性状会在家族当中得到延续,否则自然选择就不可能生效,但是他没有可靠的机制来解释遗传如何作用,尽管他确实注意到了遗传或者说“经过修改的传承”(modification by descent)的至关重要。那么在达尔文的时代最流行的遗传理论究竟是什么?已知最早的遗传学文本包括希波克拉底在公元前400年的著作,他认为生殖材料来自生物体的所有部分,因此亲代的特征会被直接传递给后代。他的证据是有一种名叫巨头族(macrocephali)的人,在孩子出生之后,父母会用手将孩子的脑袋塑造成长圆形。后来这种长圆头型就会自然而然地出现,不再需要用手捏了,就好像谢顶与蓝眼睛那样被遗传了下来。既然生殖材料来自身体的各个部分,那么自然也会来自被塑造的长圆脑袋。
五十多年后,亚里士多德提出了另一套遗传理论,质疑了希波克拉底的观点。亚里士多德不能理解,例如声音、指甲与头发之类的特征怎么能构成生殖物质,因为这几样要么没有实体,要么由死组织构成。他还注意到,儿童有时看上去更像祖父母而不是生父母。此外植物也是个大问题,像叶子这样不参与繁殖的部分怎么才能遗传下去?父母各自提供了自己的生殖物质,为什么孩子往往只能继承一方的特征?所以亚里士多德修改了希波克拉底的理论,他认为生殖物质从身体各个部分转到了生殖通路上——这是一个非常重要的理念。但是另一方面他又认为父母对于后代的贡献不一致,父亲的贡献更高一些——这倒不意外。父亲的生殖物质能够塑造胚胎,而母亲仅仅提供了所谓的“内在物质”,也就是比较无聊的部分。这种传输理论的许多变体将会在未来几百年长久流传。
唯一能与传递理论竞争的遗传理论就是预成型理论,可以追溯到圣奥古斯丁。这个理论认为当初上帝创造女性的时候已经预先塑造了她的所有后代。这个理论提出了炼金造人(Homunculus)的概念。画面上这张十七世纪的炼金造人插图出自尼古拉.哈索克(Nicolas Hartsoeker)之手,创作于1695年——根据这个炼金造人的造型,可以想见哈索克在遗传观念方面是个精子主义者。但是在达尔文时代产地理论依然占据主导地位。达尔文在1868年的论文《动物和植物在驯化环境下的差异》(The Variation of Animals and Plants under Domestication)当中讨论了泛生论。他认为有机体的有各种组织都会形成细小颗粒,他将其称作泛子(gemmule)。这种颗粒在有机体内部循环,传递给生殖细胞,从而形成有机体的各个不同部分。达尔文还认为泛子可以在休眠状态下传递给后代,他以此解释为什么亲代的有些性状在子代身上得不到表达,以及亲本的形状如何直接传给了后代。在这部1868年作品当中,他将泛生论称作“补充性假设”,这个假设对于他的自然选择理论非常重要。用他的话来说,泛生论将“目前缺乏联系的多种事实”整合在了一起。不过泛生论与希波克拉底在两千年之前提出的理论并没有显著差异。值得注意的是,希波克拉底的理论整整两千三百年来都没有遭到挑战。并不是说在此期间——尤其是在十八十九世纪——没有人做过育种试验。更准确的说法是,尽管这些这些试验并不支持经典假设,但是也提不出替代性的观点,直到孟德尔出现为止。
值得注意的是,早在孟德尔发表论文之前几十年,至少还有另外两位植物育种家发现了类似的现象。一位是1799年的英国人托马斯.安德鲁.奈特(Thomas Andrew Knight),此人将白花豌豆与紫花豌豆进行杂交,发现第二年的第一代子代都是紫花豌豆。将这些紫花豌豆进行自花授粉,在第三年得到的第二代子代既有紫花也有白花。由此他得出结论:某种“更强的趋势”导致了紫花豌豆而非白花豌豆的出现。第二位约翰.高斯(John Goss)也是英国人,1824年他更进一步,用绿色豌豆做实验给黄色豌豆授粉,发现第一代子代全都是黄色豌豆。将这些黄色豌豆再种下去,有些植株长出了全黄豌豆,有些长出了全绿豌豆,也有些长出了黄绿搭配。然后他又将这些豌豆再种下去,得到了第三代子代,发现绿色豌豆种出来的全是绿色豌豆,但是黄色豌豆的后代有些是全黄,有些是黄绿相间。那么为什么奈特与高斯没有发现遗传定律?问题在于他们没有清算豌豆的数目。就算他们数过了,也肯定没有看出这些数字有什么意义。因此这两个人都没有像孟德尔那样发现遗传机制。
四十二年之后的1866年,孟德尔再一次进行了类似实验,画面上是孟德尔亲笔书写的笔记插页,其中详细记述了实验数据,正是这些数据让孟德尔得以在灵光一闪之下提出全新的假设。孟德尔的伟大之处很容易就被豌豆育种的晦涩事实与数字所掩盖,但是魔鬼隐藏在细节当中,伟大只有通过看似平庸的事实才能得到揭示。就像奈特一样,孟德尔也用紫花与白花豌豆做实验,也注意到第一代子代F1全是紫花。然后他也用这批豌豆自花授粉,得到了既有紫花也有白花的后代F2。但是孟德尔清算了F2当中紫花与白花的数量,发现在总计929株F2当中有705株紫花,224株白花。大家或许会注意到,这两个数字接近总数的3/4与1/4。孟德尔将紫花性状——出现在所有F1与3/4的F2当中——称作显性,将白花性状称为隐性。换句话说遗传是颗粒性的,并不是将亲本性状像油彩那样混合在一起。
另一方面,孟德尔也进行了黄色与绿色豌豆的杂交,确定了黄色是显性性状,绿色是隐性性状。在8023颗F2豌豆当中,6022颗是黄色,2001颗是绿色,依然接近3/4与1/4,或者说三比一。孟德尔确定了高斯的观察结果,但是他还更进了一大步。他发现在519个黄色豌豆豆荚当中,166个全都是黄色豌豆,353个夹杂着绿色,而且黄色与绿色的比例同样是三比一。孟德尔意识到,166个纯种黄色豌豆荚与353个混种黄色豌豆荚的比率很接近一比二。换句话说,F2当中三个显性对一个隐性的表面比率实际上意味着一个纯种显性加上两个混种显性再加上一个纯种隐性——最后这组豌豆的后代永远全都是绿色的。他将这个实验进行了好几代,发现纯种豌豆总会保持纯洁,而混种豌豆每一代都会产生一比二比一的比例。孟德尔对待数据的态度很谨慎,他又对比了七对其他性状,发现每一对性状都分为显性与隐性,F2后代总会产生三比一的比例,或者说伪装之下的一比二比一。
这一切究竟意味着什么?孟德尔提出了一项假设来解释自己的数据,他用Y表示显性,用y表示隐性。在他的实验当中,黄豌豆是Y,绿豌豆是y。他正在考虑怎样的因素导致了这些性状的显现。按照孟德尔的假设,性状并不是按照经典理论从上一代直接传到下一代。遗传因子其实是某种相互离散的颗粒,每一种颗粒只会负责特定的性状。就每个形状而言,每个个体都会从亲本双方各自获得一个颗粒,而且这两个颗粒在产生生殖细胞的时候相互分离。比方说亲本是YY的黄豌豆与yy的绿豌豆,那么F1后代就全都是Yy,这批豌豆全都是黄色的,因为黄色是显性性状。进一步说,这批豌豆自授粉之后会产生YY、Yy、Yy与yy的F2后代,黄色对绿色是三比一——如果你考虑到遗传因子本身的话,实际上是遭到伪装的一比二比一。另一项事实也没有逃过孟德尔的注意:携带Y的卵细胞被Y或y划分授粉的可能性一样高,换句话说,卵细胞与花粉——就动物来说则是卵子与精子——的受精结果是随机的,无论它们携带什么因子。这就是孟德尔的独立分配定律。
孟德尔走到了意义无法估量的革命前沿,但是谁都没有注意到。他的论文在1866年发表,比达尔文的泛生论论文早了两年,届时达尔文将会提出基于泛子的遗传特征混合理论,而泛子又来自有机体的一切部位。站在我们今天这个即时沟通手段业已完善的时代回头来看,令人难以置信的是达尔文似乎从没阅读过孟德尔的论文。如果达尔文遇到了孟德尔并且讨论了孟德尔的实验结果会怎么样?达尔文肯定会感到摸不着头脑,不明白为什么孟德尔的F1豌豆不是黄绿色。孟德尔的实验表明遗传特征的模式肯定是颗粒状的,我真想知道达尔文对此有什么说法。
孟德尔的论文几乎遭到了彻头彻尾的忽视,三十四年之后的1900年才被被三名科学家各自重新发现。他们分别是奥地利的冯.丘歇玛克-赛塞内格(Armin von Tschermak-Seysenegg),荷兰的H.德弗里斯(H·Devries)与德国的C.科伦斯(C·Correns)。这三个人当时正在研究各不相同的植物种类。科伦斯率先提出了孟德尔定律(Mendel Regel)的说法来形容遗传基本原则:决定相对性状的离散颗粒状遗传因子会在生殖细胞——或者说配子——形成时分离开来,又会在受精时重新聚合。德弗里斯提出了变异一词来形容生物体突然出现的变化,变异提供了自然选择的原材料。威廉.贝特森(William Bateson)与伊迪丝.瑞贝卡.桑德斯(Edith Rebecca Saunders)在1902年率先将孟德尔定律运用在了动物身上。他们研究家鸡的时候发现,与正常的鸡爪相比,额外多一个脚趾是显性性状。
贝特森与桑德斯还根据孟德尔定律检视了许多未能得到解释的早期数据,甚至重新考察了达尔文于1868年发表的狮子花研究成果。狮子花的一种形态是中心对称的反常整齐形态,与之相对的则是常见的不规则上下双唇形态。达尔文认为他的杂交试验表明产生正常花朵的倾向在F1代呈压倒性趋势,但是“过了一代之后,反常整齐倾向似乎变得越发强大起来”。根据孟德尔的理论,我们可以推测反常整齐形态是隐性性状,所以会在F1代消失,在F2代以一比三的比例出现。由此可见,经典遗传理论与孟德尔理论之间可谓泾渭分明。根据经典理论,性状直接从上一代的花朵部位传到了下一代的对应部位。但是根据孟德尔理论,遗传是非直接的,通过颗粒化的遗传因子来完成。正是这些遗传因子从一代传到另一代。无论是花的性状还是其他什么性状之所以得以发展,仅仅是因为受精时接受了一套随机的遗传因子组合。双亲各自贡献的遗传因子都维持了自身的完整性,不会与另一方混合。更重要的是,遗传因子在有机体体内的存在并不会使其遭到修改。换句话说,经典理论当中希波克拉底提出的被拉长的脑袋可以遗传的案例根本不可能发生。
在孟德尔的成果被重新发现之后,二十世纪初生物学的最主要问题在于这套理论与进化的关系不清不楚,人们为此展开了激烈辩论。野生种群的性状差异看上去是连续性的,更像是混合遗传物质的模型,而不是孟德尔的离散模型。而且孟德尔又要怎样解释新物种出现这种跨度极大的事件?解决这个问题的关键在于研究野外种群,我们现在将这个领域称作种群基因学。这方面的关键人物之一名叫罗纳德.费舍(Ronald Fisher),他开创了所谓的现代综合论。1918年他发表了开创性的论文《孟德尔遗传假设下的亲族之间的相关性》。这个题目听上去很乏味,但是远非如此。费舍在论文中表明,连续性的差异可以由许多离散的决定因素所导致——如今我们将这些因素称作基因座——因此孟德尔的遗传理论与自然选择驱动的进化完全相符合。到了二十世纪二十年代,孟德尔原则在种群方面的应用在费舍、J.B.S.霍尔丹(J.B.S.Haldane)、休厄尔.赖特(Sewall Wright)、多勃赞斯基、E.B.福特(E. B. Ford)、恩斯特.麦尔(Ernst Mayr)等人的推动下得到了更进一步的发展。可惜今天时间不足,容不得我详细讲述这几位在其他方面的的重要贡献,例如维护基因多样性,研究基因多态性,以及确定新物种产生生殖隔离的机制。不管怎么说,关于种群基因学的探索在我看来是暨1866年孟德尔提出遗传理论并且在二十世纪初得到重新发现之后的第二场生物学基因革命,这第二场革命带来了生物学的大一统理论:基于自然选择的进化范式整合了遗传机制,可以适用于种群层面。基因层面的变化也就是变异提供了自然选择的原材料,从而在世代延续当中塑造进化。
接下来的第三场革命是染色体理论的问世。决定因素与颗粒什么的说着好听,但是它们究竟是什么?究竟位于哪里?“在哪里”这个问题由托马斯.亨特.摩根(Thomas Hunt Morgan)给出了回答,大约与现代综合论的发展同步。至于“是什么”这个问题还得再等几年。巧合的是,托马斯.亨特.摩根于1866年生于肯塔基州,正是孟德尔发表论文的那一年。当摩根在1910年开始研究果蝇的时候,孟德尔的发现已经被总结成了两大定律。刚才我们已经谈到了第一定律,即分离定律。第二定律名为自由组合定律,今天我们称之为独立分配定律。这条定律意味着当新一代诞生时,不同的遗传因素会形成新的组合并且相互独立。孟德尔将这个现象称作二元杂交(dihybrid cross)。比方说,一株高杆紫花植物与矮杆白色花植物相互授粉,那么紫花与白花因子的遗传将会独立于高杆与矮杆因子的遗传,因此下一代的高杆、矮杆、紫花、白花各种组合的比例完全可以预测。但是摩根的天才在于他发现了第二定律与现实的不相符之处,并且足够自信地提出了解释。一开始摩根将紫眼短翅的果蝇与红眼长翅果蝇杂交,他用pr代表紫眼,vg代表短翅,用pr+与vg+代表红眼长翅——+在基因学当中意味着正常。根据孟德尔第二定律,孟德尔的杂交培育出紫眼长翅果蝇与红眼长翅果蝇的频率应该一样,因为遗传因子应当独立分配,但是事实并非如此,紫眼长翅的杂交果蝇非常少见。于是摩根向前跃进了一大步,他认为眼睛的颜色与翅膀的尺寸一定相互关联,他将这个现象称作连锁(linkage)。摩根通过果蝇杂交发现了很多这样的例子,但是关键在于连锁关系仅仅存在于四因子组合当中。
正当摩根进行果蝇杂交的同时,十九世纪末二十世纪初的早期细胞生物学家也很忙碌。摩根注意到了这些人的工作。例如奥斯卡·赫特威格(Oscar Hertwig)于1875年研究海胆的时候发现了受孕机制;奥古斯特.魏斯曼(August Weismann)于十九世纪八十年代通过显微镜区分了遗传物质——他称之为种质(germ plasm)或者种系(germline),也就是组成配子的物质——与生物体的其余部分或者说胞体(soma)。由此他得出结论:雄性与雌性生殖细胞的细胞核一定是遗传特质的携带者。接下来几位新兴细胞生物学的信徒在细胞里发现了线状结构,将其称之为染色体。1888年,海因里希.冯.瓦尔德耶-哈兹(Heinrich von Waldeyer-Hartz)首次观察到了这一结构。画面上这张染色体手绘图出自瓦瑟尔.弗莱明(Walther Flemming)之手,他的观察对象是蠓虫的唾液腺细胞。但是谁也不知道这些线状物体究竟有什么作用。
当孟德尔的成果在二十世纪初得到重新发现之后,西奥多.鲍威里(Theodor Boveri)和沃尔特.萨顿(Walter Sutton)各自提出,可以用染色体来解释遗传因子在新个体形成时的分布情况,根据则在于生殖细胞内部染色体数量在受孕之前与之后的比例。通过染色体,遗传因子可以结合也可以分离,因为体细胞内部的染色体数量是生殖细胞的一倍。
现在请大家看一段影片,这是我最喜欢的影片,拍摄对象是我正在研究的秀丽隐杆线虫的胚胎。可以看到,雄性与雌性生殖细胞的细胞核即将融合在一起,这一事件标志着新生命的开始,之后受精卵将会开始细胞分裂等一系列进程。我总觉得在显微镜上看到这一幕非常惊人。配子包含着来自父本的一半遗传物质和来自母本的一半遗传物质,两者结合再次拼成整体。染色体在生成生殖细胞或者说配子的时候数量减半的现象叫做减数分裂(meiosis)。鲍威里和萨顿认为,染色体会在配子形成时重新分配遗传因子,这一理念有力应和了孟德尔第一定律即分离定律。
理论基础已经打下,就等着摩根将细胞生物学与遗传学联系在一起。事实证明,染色体正是将众多遗传因子链接在一起的连锁群。而遗传因子组合或者说连锁群的数量也就是细胞里的染色体数量——就果蝇来说是4条,就人类而言是23条。摩根发现的连锁现象完全证明且显著扩展了鲍威里和萨顿的染色体遗传理论,同时又在很大程度上限制了孟德尔第二定律,因为这条定律只能用于位于不同染色体的遗传因子。摩根发表了他的染色体地图,在染色体上标注了不同的遗传因子,就好像是项链上的珠子。但是摩根还更进一步发现,他的连锁理论依然还有不符合实际之处。尽管有些遗传因子位于同一个颜色体上,但是依然会出现一定程度的自由组合,尽管十分罕见。例如摩根的杂交确实产生了不多的几只紫眼长翅果蝇,摩根将这个现象称作染色体交换(crossing over),并且构想了不同的染色体之间怎样在交叉点(chiasmata)交换组成部分。我们可以在细胞当中观察到这种现象,例如画面上的蚂蚱睾丸细胞——我很想知道人们怎么想到观察这些奇怪的生物系统,总之我们可以看到染色体在蚂蚱体内的交叉点。交叉理论一度深受怀疑,直到1931年2月,哈莉特.克雷顿(Harriet Creighton)与芭芭拉.麦克林托克(Barbara McClintock)以更高的分辨率观察到了染色体相互交换构成部分。现在我们将这一现象称之为基因互换重组,这是在生殖期间产生基因多样性的重要机制。
因此第三场革命就是染色体遗传理论,这一理论链接了孟德尔遗传定律与染色体的物理结构。染色体符合孟德尔第一定律,但是当遗传因子位于同一个染色体上的时候又会违背第二定律。摩根的工作完全非同寻常,谁能想到在只有一个微米这么长——也就是1/1000毫米——的染色体上居然有可能确定上千个遗传遗传因子,能够概括一切有机体的一切特质,而且摩根仅仅通过研究果蝇就取得了此等发现?摩根在1937年荣获诺贝尔奖的时候,他的工作被比作天文学家通过计算寻找尚未被观测到的天体,后来又通过天文望远镜发现了这些天体。但是实际情况远不止如此,摩根的预测远远超过了这种程度,因为他的发现意味着原则上的全新事物,从没被观察过的事物。用摩根自己的话来说,“遗传的根本方面居然如此简单,这使得我们有理由期望自然界或许完全可以得到理解。她那交口传扬的不可解特质再一次被证明只是幻觉,只是因为我们的无知才得以维系。这一点着实令人振奋。如果我们的世界真像我们的某些朋友想象的那样复杂,我们或许会绝望地认定生物学毕竟无法成为一门精准的科学。”
基因这个词最早于1909年由威廉.约翰森(Wilhelm Johannsen)发明,用来形容遗传的实体与功能单位。基因/gene这个词取自达尔文的泛生论/pangenesis。但是基因的角色直到二十世纪四十年代才通过乔治.比德尔(George Beadle)与爱德华.塔特姆(Edward Tatum)的里程碑式研究得到确定,我们由此来到了第四场革命的起点,也就是分子生物学革命。这场大规模革命其中又包含好几场次级革命,其中有些比另一些更出名。为了长话短说节省时间,我总结了接下来四十年的主要成果,并且将其划分成了好几场次级革命。接下来这段走马观花式的简介将会让我们一路抵达二十世纪八十年代。
比德尔与塔特姆研究的对象是名为脉孢菌的单倍体真菌。他们用辐射处理脉孢菌致使其变异,然后寻找有趣的变异表型。他们发现很多变异真菌都营养不良,因为它们合成精氨酸的路径出了问题。比德尔与塔特姆设法在不同阶段阻断变异体的合成路径,然后发现每一种基因控制生化路径上的一种酶,由此他们提出了一基因一酶假说。他们的工作提供了意义显著的大一统理念,在基因学与生物化学这两大重要研究领域之间搭建了桥梁。所以第四场革命当中的第一场次级革命就是一基因一酶/一蛋白质理论的问世。
那么基因由什么构成?弗雷德里克.格里菲斯(Frederick Griffith)在1928年率先回答了这个问题。他的研究对象是肺炎链球菌。有些细菌如果注射到老鼠体内就会致命,另一些毒性较弱的菌株则不会致死。格里菲斯发现了一个令人困惑不解的现象:假如将非致命菌株与高温灭活的致命菌株——其本身并不致命——一起注射进老鼠体内,老鼠还是会死。从死老鼠身上提取到的菌株注射到活老鼠身上依然致命。显然,灭活致命细菌的残骸致使非致命细菌转化成了致命细菌。换句话说细菌的细胞被转化了。快进到1944年,奥斯瓦尔德.埃弗里(Oswald Avery)与两位同事科林.麦克劳德(Colin MacLeod)以及麦克林.麦卡锡(Maclyn McCarty)试图用化学方法逐一破坏死细胞里的所有主要化学物质,从而确定破坏哪种物质会让死细胞失去转化活细胞的能力。想必大家都能猜到他们发现了什么:DNA。既不是多聚糖或者蛋白质,也不是脂肪或者RNA,就是DNA,这就是导致转化的物质。他们首次证明了基因由DNA构成。这是第二场次级革命:DNA是基因的物料。
老实说这一点有些令人有些失望。复杂程度这么低的分子怎么能够为地球上所有复杂生物编码呢?DNA早已为人所知,因为早在1869年弗雷德里希.米歇尔(Friedrich Miescher)就将它分离了出来。他从诊所的脓液当中提取出了DNA,因为脓液里边有很多白细胞,而白细胞的细胞核特别大。然后俄国生物化学家菲巴斯.利文(Phoebus Levene)在1919年发现DNA是多聚核苷酸。就像所有化学家一样,他也喜欢分解物质——这次他用得是水解法。结果他发现核苷酸的成分是四种核苷酸碱之一、一个糖分子以及一个磷酸基团构成。这就是第三场次级革命:DNA的成分是四种核苷酸碱、一个糖分子以及一个磷酸基团。利文认为这些成分构成了他所谓的四核苷酸(tetranucleotide)结构,其中不同核苷酸分子按照固定顺序排列:G-C-T-A-G-C-T-A。但是科学家们最终意识到这样的结构过于简单。核苷酸的分布顺序实际上多种多样。第四场次级革命:DNA大分子里的碱基顺序可以改变,这样的排序或许可以充当编码体系。
接下来,埃尔文.查戈夫(Erwin Chargaff)发现腺嘌呤/A的数量大致等同于胸腺嘧啶/T,鸟嘌呤/G大致等同于胞嘧啶/C。查戈夫在很大程度上受到了1944年埃弗里论文的启发。他写道:“埃弗里给了我们一种新语言的第一套文本,或者说他向我们指明了去哪里寻找这套文本。我决心一定要找到。”这就是第五场次级革命:A等于T,C等于G。
查戈夫的工作,以及英国研究人员罗莎琳德.富兰克林(Rosalind Franklin)与莫里斯.威尔金斯(Maurice Wilkins)非常重要的X光晶体成像成果,为弗朗西斯.克里克和詹姆斯.沃森在1953年确定DNA分子的三维双螺旋结构铺平了道路。在《自然》期刊发表的论文上,沃森与克里克写道:“我们拟提出脱氧核糖核酸(DNA)盐的一种结构。这一结构拥有一些崭新的特征与重要的生物学意义。”这项成果的意义极其重大。两万五千股DNA可以塞进一根人头发的宽度里。这是第六场次级革命,也是特别重要的一场:确定DNA的结构为一根双螺旋长分子链,精巧、简单、有序、规则。每一条螺旋都由糖与磷酸盐都成的强健脊梁构成,碱基位于内侧,A与T配对,C与G配对。
有些人认为,沃森与克里克确定的DNA结构是二十世纪最重要的生物学发现。这么说是因为双螺旋结构满足了对于遗传物质来说的三个要求。首先是有序。四个碱基构成了四字母的字母表,借此写成的基因编码可以确定构成蛋白质的氨基酸的顺序,而蛋白质对于细胞功能至关重要。弗雷德里克.桑格(Frederick Sanger)在1951年首次确定了胰岛素蛋白质的氨基酸顺序。氨基酸排序非常需要基因代码,或许基因代码可以通过核酸排序在DNA当中写下信息,然后将这些信息翻译成蛋白质里的氨基酸排序。其次,如果DNA碱基的顺序确定了蛋白质的氨基酸顺序,而突变又有可能将一种碱基变成另一种碱基,那么突变就可以改变蛋白质,从而改变细胞功能,这就很了不起了。基因多样性是孟德尔的研究课题与达尔文进化的驱动力,现在我们找到了这一点的根本原因。难怪克里克会在1953年2月28日冲进剑桥大学老鹰酒馆高呼道:“我们发现了生命的奥秘!”第三是复制。沃森与克里克在论文结尾写道:“我们注意到,我们假设的配对方式显然意味着基因物质可能的复制机制。”如果核苷酸可以相互配对,而且细胞内部存在自由的核苷酸与相应的酶,DNA就可以复制自身。以下是一项了不起的事实。每一天我们全身会产生600亿个新细胞,在这一过程当中得到复制的DNA总共有一千亿米长,足够从地球到火星拉个来回。
所以DNA是编码,这种编码包括了构成一切生命的所有信息,是构建生命的指导手册,无论这生命是线虫、猫、苍蝇、人类还是恐龙。它的规整性与可预测性使其得以存储构建生命所需的大量信息。这些信息会传输进有机体的每个细胞,因为在每个细胞分裂之前DNA都完成了完美的复制。同样,所有的信息也都会根据孟德尔定律传入每一个生殖细胞或者说配子,传入种系,然后流传到每一代。
第七场次级革命是解码。证明密码子(codon)——也就是对应一种氨基酸的核苷酸组合——包含三个字母的过硬证据来自1961年的一场极其优雅的实验。弗朗西斯.克里克、西德尼.布伦纳(Sydney Brenner)与同事们研究了专门感染细菌的噬菌体病毒的变异情况。他们发现,如果一次为噬菌体的DNA添加或者减少三个核苷酸,病毒并无大碍,可见DNA代码的基本单位由三个核苷酸组成。还是在这一年,马歇尔.尼伦伯格(Marshall Nirenberg)、J.海因里希.特伊(J.Heinrich Matthaei)以及同事们破解了密码子,将每一种氨基酸与特定密码子联系在了一起。
接下来让分子生物学家挠头的问题是基因调控的机制,这就是第八场次级革命。问题在于,假如DNA编码赋予了细胞特定的特质,而且生物体当中的所有细胞都包含同样的DNA序列——因为DNA序列定义了这个物种——那么为什么细胞能在不同的时间、在身体的不同部位做出不同表现?这项突破要归功于一位年轻的巴黎分子生物学家雅克.莫诺(Jacques Monod)与一位打算从事外科医生的医学生弗朗索瓦.雅各布(Francois Jacob)。纳粹入侵法国之后,雅各布加入自由法国运动成为了一名军医,后来身负重伤。莫诺加入了法国本土的抵抗组织,并且在组织里遇到了另一位分子生物学家安德列.利沃夫(André Lwoff)。巴黎光复之后莫诺加入了法军,结果在美军的一座战地流动图书馆里发现了艾弗里主张DNA是遗传物质的论文——想想这一幕吧,战场上的军人居然在空闲时间阅读分子生物学文献——这使得他重新燃起了对于基因学的兴趣。战争结束后,莫诺与利沃夫开始合作研究噬菌体与细菌。然后他们又拉上了雅各布,他因为受伤太重只得放弃了外科手术这一行。接下来十年里,这三个人成为了基因学史上最富创造力的研究组合。他们发现了基因开启与关闭的机制,换句话说同样的基因物质在不同的细胞里可以得到不同的解读,产生不同的蛋白质。最后他们还发现某些基因形成的蛋白质可以与DNA结合,从而阻断其他基因的表达。他们的研究成果完全是通过基因证据推导出来的,而这些证据则来自对于变异效果的研究。
我强烈向大家推荐贺瑞斯.弗里兰.贾德森(Horace Freeland Judson)的《创世第八天》,他在书中描述了分子生物学革命的酝酿阶段。在他看来,莫诺等人的成果“使得原本完全黑暗的东西变得非常简单”。基因开关的概念远远不仅只能应用于细菌的酶与病毒。他们理解了、并且极其雄辩地阐述了他们关于基因调控的发现可以怎样解答关于细胞分化以及胚胎发育的奥秘——所有细胞都具有同一套基因的问题在这两方面格外突出。“对大肠杆菌成立的规则对于大象也同样成立。”我们就这样来到了分子生物学革命的最终阶段,多年成果积累的高峰。我们在分子层面理解了基因,包括DNA在生物体内部以及代际之间的复制机制,这意味着遗传物质的流传可以以更切实的方式得到理解。这样的理解涉及真正的分子,而这些分子的表现又完全与它们的结构相一致。大一统的基因进化论就此得到了分子层面的理解。
那么接下来往何处去?接下来的第五场革命是正向遗传学的问世,这也是我非常看重的一场革命,通过基因,我们不仅理解了遗传机制,还理解了很多其他生物学问题。赫尔曼.穆勒(Hermann Muller)在1927年研究果蝇的时候发现,用X光或者特定的化学物质来处理有机体可以导致大量可遗传变异,显著影响有机体的形态或者说表型。这当然是因为变异改变了DNA序列,阻碍了对应变异DNA编码的特定蛋白质发挥作用。生物学家开始运用基因来研究变异,试图理解基因如何影响了其他在当时依然神秘的生物学现象。这其中最重要的现象就是生长:一个有机体怎样从卵子长成成体?成体又要怎么确定自身的正确尺寸?细胞如何确定自己以正确的数量出现在正确的时间、正确的位置并且发挥了正确的功能?这看上去是个非常复杂的问题,但是这个问题可以借助模型有机体来理解,例如果蝇。其他登场较晚的生物还有在二十世纪七十年代得到使用的秀丽隐杆线虫。只要看看实验生物的变异体呈现出了怎样的表型,就可以看出这种做法多么有用。例如画面上是一个变异果蝇的的头部,在该长触角的地方长出了一条腿——我认为弄错这种事还挺不容易的。这个实验表明,导致这种缺陷的单独一个变异基因扮演着非常重要的角色,负责让腿长在正确的位置,避免触须长在错误的位置。通过观察各种错误,我们就可以确定正确的过程通常是怎么样子。
1980年,克里斯汀.纽斯林-沃尔哈德(Christiane Nüsslein-Volhard)与艾瑞克.威斯乔斯(Eric Wieschaus)共同发表了一篇论文,两人最终获得了诺贝尔奖。他们收集了大量有趣的果蝇变异,揭示了果蝇有机体不同的生长方面在分子层面上如何运作。接下来西德尼.布伦纳在1973年将我最喜欢的秀丽隐杆线虫推上了前台。画面上半部分是正常的虫子,下半部分则是两种变异体,一种变得更短更粗,另一种变得更细更长。这些虫子表明了动物的尺寸如何得到调节,不同的动物怎样确定自己的特定尺寸。利用变异使得确定尺寸的过程出错,我们就能理解这个过程在正常情况下怎么运作。
这种研究方法自从二十世纪二十年代就开始得到运用,到了二十世纪七八十年代更是焕发了新的活力,因为这时生物学家们理解了基因的本质,可以利用摩根发现的连锁现象与染色体理论来标注基因变异。其次,这时我们掌握了确定DNA序列的技术。第三,这时我们可以制造转基因动物。这三条意味着我们可以确定指出出错基因的位置,在体外将DNA分离出来,折腾一番之后再引入苍蝇、线虫、酵母菌甚至老鼠体内。这种能力引爆了一大片生物学领域。现在我们在分子层面上相当详细地理解了细胞分裂如何被控制,胚胎细胞如何确定自己的正确身份,细胞怎样迁徙到正确的位置,性别在分子层面如何被确定,为什么有些细胞会遭受程序性死亡……我可以花好几个小时来举例子。更进一步,转基因技术让我们可以将一个物种的基因引入另一个物种。保罗.纳斯(Paul Nurse)在二十世纪八十年代率先推行了这项技术,揭示了人类体内有一个基因可以对应酵母菌所具有的cdc2基因,这个人类基因可以用来救活那些原本因为cdc2基因缺陷而无法正常生长的酵母菌。换句话说,人类及与酵母菌基因非常类似,它们的功能保持不变,这一点非常重要,因为这意味着就像我们之前怀疑的那样,研究简单有机体可以增加我们对于人类生理与疾病的理解。模式生物由此具备了重大的生物学意义。
利用基因方式,我们还可以理解寿命长短如何得到预先确定。画面上可以看到正常线虫的寿命分布,经历了daf2基因变异之后,线虫的寿命就远远超过了正常程度。我们甚至正在开始理解基因如何控制行为。画面左边是正常的线虫,右边是变异的线虫——更准确地说,右边也不是完全的变异体,因为这里所谓的变异是天然发生的而不是被化学手段诱发的。左边这些线虫来自澳大利亚,他们喜欢聚集成群;来自英国的线虫看起来似乎更加孤僻。这表明我们可以借助基因来理解行为,因为这两组线虫之间的唯一区别就是一个基因的变异。正向遗传学的最为革命之处在于并无偏见。研究人员观察一批变异个体,直到发现某个个体显现出特定的缺陷,由此揭露重要的生理机制,并且通过确定变异基因的序列在生物化学层面理解这一机制。这样的研究之所以没有偏见是因为我们事先并不会针对得到研究的生物化学过程做出任何假设。基因学家可以通过表型分析找到重要的基因,却不能挑剔他们想要研究什么基因什么蛋白质,因为基因会将他们引向任何地方。他们的研究完全以表型为导向。我们在过去四十年取得了极大的进步,基因帮助我们深入理解了生命的原理。这些非同寻常的努力就是我所说的第五场革命:利用变异来理解一切源自基因编程的生物学现象——基本上就是一切生物学现象。
第六场也是倒数第二场革命是基因组测序。弗雷德里克.桑格(Frederick Sanger)于1977年开始了这项工作。桑格早在二十世纪五十年代就发现了为蛋白质进行氨基酸测序的方法,然后他又研究了DNA测序。于是他拿到了两个诺贝尔奖。针对小段DNA测序的技术问世之后,下一步就是提升规模,针对整个基因组进行测序。1995年,流感嗜血杆菌成为了最早得到基因组测序的非共生生物,接下来是1996年的酵母菌。1998年,秀丽隐杆线虫成为了第一种得到测序的多细胞生物,果蝇测序在2000年完成。人类基因组测序在2000年完成草图,2003年大功告成。这是了不起的成就,几百名世界各地的研究人员花了十三年时间与二十七亿美元才完成这项壮举。如果你想以每分钟二百个字母的速度输出整套人类基因编码,需要不停歇地打字二十九年。但是我们每个人体内的四十万亿个细胞当中的每一个细胞都包含这一套编码,我们的酶机器只用几分钟就能复制一套。
约翰.萨尔斯顿爵士(Sir John Sulston)是人类基因组测序项目英国部分的领头人,并且一直在孜孜不倦地确保这些数据处在公有领域。用他的话来说:“我们在人类历史上走到了当下地步,有史以来我们手里第一次掌握了创造人类的全套指南。”从那以后的进展更加非凡。科技发展如此之快,以至于只用几个小时与不到一千美元的投入就可以完成人类基因组测序。上千种有机体都已经接受了测序。海啸一般的大量数据就连存储都成了问题,更不用说分析了。2013年有人估算,全世界的生物学家每天会产生十三Pb的数据——一Pb等于一百万G。用DVD的存储的话,全摞起来有2.2英里高。至于今天我们产生数据的速度比起那时候还要快得多。
所以这第六场革命为我们带来了很大的挑战。我们很容易就会因为基因组信息的可能用处而目眩神迷。针对你的癌细胞进行基因组测序或许可以让肿瘤学家订制专属于你的治疗方案从而增加成功率。我们猜测医疗人员早晚会使用基因组信息来预测一个人可能得什么病,并且试图最小化甚至消除风险,为人们提供个人化的预防药物。实际情况并非如此直接了当。因为人类疾病往往并非由容易追踪的单一基因变异导致——这种疾病的典型例子是囊性纤维变性——大多数疾病例如心脏病、癌症或者精神疾病都源自许多不同基因与彼此以及环境之间的复杂互动。公众往往错误地认为科学家发现了“导致某某疾病”的基因,更不用说导致某某特征的基因了,例如酗酒、健忘、智商或者美貌。人类生理与基因之间很少存在如此决定性的关系。基因最好被视为概率因素,大量基因都会影响可能性,但是最终结果在很大程度上是基因、生理与环境影响共同作用的结果。因此我并不认为我们非得对于基因医学抱有过于敌托邦的惨淡观点。伦理争论确实存在,例如单基因失调。此外迟发性疾病也是个问题:我们想不想提前知道自己老年可能患什么病?如果你想知道,你的家人该怎么办?你的诊断结果会不会影响他们?如果他们不想知道又该怎么办?诊断结果带来的心理冲击应当怎么应对?会不会造成基因歧视?
谢天谢地的是,根本不可能仅仅依靠简单的基因测试就确定某人的智商或者创造力,或者此人患上心脏病、糖尿病、精神分裂症的可能性。这其中的因素过于复杂了。我敢肯定,将会有雨后春笋一般众多的互联网公司提供此类测试服务,他们或许应该遭到禁止。并不是因为它们会给你提供糟糕的信息,而是因为它们几乎一定会给你提供误导性的信息。另一方面,概率性观点会告诉你怎样避免个人风险,具体做法或许涉及少吃脂肪,少喝酒,多锻炼——我从哪里听过这些说法?我不认为基因学不会构成任何社会伦理挑战,可能出现的问题包括基因信息的所有权、隐私权以及访问权,这些都需要辩论。辩论必须发生在充分知情的利益相关方之间,而不是遭到另有打算的游说集团左右。
以上就是第六场革命,既基因组革命。这场革命的结果还不清楚,但是可以肯定,基因组测序信息让所有生命变得更加紧密了。画面上是现代版本的达尔文生命之树。我们已经掌握了上千种基因组信息,通过比较我们可以以前所未有的清晰程度研究不同物种之间的进化关系。DNA不仅仅是制造有机体的手册,也是生物进化的历史书,纪录了让我们成为我们、让一切生物成为如今模样的一切偶然基因变异。我们全都通过万万亿份不断复制的基因在四十亿年间联系在一起。想一想吧,假如达尔文与孟德尔知道了这一切,他们会多么兴奋?
我要介绍的最后一场革命,也就是第七场革命,让我们走向了未来。但是这场革命与第六场革命不同,眼下才刚刚开始。我说的就是借助CRISPR-Cas9进行基因组编辑。基因学家发现了几种技术,可以靶向改变核苷酸顺序,从而编辑或者改变有机体的基因。但是CRISPR的革命之处在于效率极其高而且用途广泛。这项技术利用了细菌的天然免疫系统当中的分子剪刀,可以去掉一段DNA,例如由病毒传播的DNA。这些分子剪刀可以极其精确地切掉一段DNA,然后通过合成制造的向导RNA进行改造。通过将向导RNA与对应的基因组序列杂交,我们可以确定剪切的位置。细胞修复剪切破损时候会出现错误,从而将被剪切的基因永远排斥出去。又或者可以趁机向这段基因组序列引入额外的修改。因此这项技术具有重写基因编码的潜力,通过修改受影响细胞的基因,就像擦掉笔误那样,我们或许可以解决基因疾病。此外我们还有可能针对一系列疾病研发激进的全新疗法。尽管单一基因失调疾病很少见,但是将各种罕见病加起来依然会影响很多人,所以整体来说它们并不少见。英国基因同盟组织估计5.5%的英国人会在二十五岁之前患上六千多种孟德尔病——既单一基因变异导致的疾病——当中的一种,总数就是350万人,基因编辑对这些病人及其家人的影响将会非常大。人类胚胎也可以被修改,导致疾病的变异可以被抹掉。这样的治疗不仅影响个人,还会影响此人的子子孙孙。
这样做真的没事吗?研究机构、研究资助人与科学家号召人们针对修改人类基因的伦理问题展开全新辩论,赶在科学进度大幅超越公众意见之前。科学家的主流意见认为这项手段过于新颖,目前不可能知道其中危险何在。他们担心公众反对会妨碍更新更安全疗法的研发。因此我们是否应当勇敢面对这一挑战?用修昔底德的名言来说,这项挑战可谓“荣耀与危险共存”。这样做是不是在扮演上帝?修改人类种系是不是决不能越过的界限?这是不是通向所谓“设计婴儿”的第一步?无论我们如何回答这些问题,有一件事非常清楚:公开讨论是关,就像基因组诊断技术一样。公共辩论必须在高层次展开,各方都要理解科学,从而避免话术替代事实。公众对于科学的信任取决于公开、透明、有效的监管,必须采取这样的做法。科学无法回答“怎样正确应对技术的潜在用途”这个问题,必须由全社会来给出答案。对于我来说CRISPR的神奇之处在于进化再一次向我们展现了前进的道路。进化创造了这么多神奇的酶,而我们则有幸足够聪明来使用它们。
转了一圈之后我们又回到了起点,从达尔文缺乏遗传理论的窘境到孟德尔,从摩根到一代代DNA研究人员,从基因组测序到基因组编辑,这是一场走马观花之旅,我遗漏了无数重要的革命,例如芭芭拉.麦克林托克(Barbara McClintock)对于跳跃基因或者说转座基因的研究,表明基因组并非静态,而是高度动态的存在;我略过了很多技术革命,例如为基因组编辑铺平道路的PCR与克隆技术;我没时间介绍最新的理念,例如表观遗传以及RNA革命——实际上基因领域充满了令人兴奋的革命,一个小时根本说不完。如果我要用一句话总结我所说的一切,那就是要注意生物学内部非凡的对立统一,一方面是复杂得惊人的多样性,一方面是稳定得惊人的根本机制。我想大家都会同意,生物学革命正在逐渐加速,我们生活在令人兴奋的时代。你要问J.B.S.霍尔丹生物学有没有未来,他会这么说:“在预测科学研究的未来时应当注意一条一般性定律:预期之外的情况总会发生。所以我们可以确定,未来肯定会使得任何详细预言看上去很蠢。尽管研究人员总会比一般人看得更远,但是像我这样一个对于植物学完全无知的人要是以为自己居然可以挂一漏万地概括整个生物学领域,那实在是自以为是至极。”我希望大家也能够原谅我的挂一漏万。最后请容许我再引用一句霍尔丹的名言:“我怀疑宇宙不仅比我们想象的更奇怪,甚至比我们能够想象的更奇怪。”谢谢大家。
- 相关回复 上下关系2
🙂233-Alison Woollard:生物学的七场基因革命
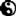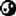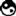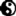
🙂补 8 万年看客 字15301 2023-01-17 07:28:28